1. Introduction
Cleft palate is one of the most common birth deficits in humans. It is considered a multifactorial disease and both genetic and environmental factors play a role in its development (Murray and Schutte,
2004). In our previous study, we have established that 2,3,7,8-tetrachlorodibenzo-
p-dioxin (TCDD) and dexamethasone (DEX) induced cleft palate in mice (Chai et al.,
2010). However, the molecular mechanisms of TCDD and DEX induced cleft palate were largely unknown. Vitamin B
12 is one of the diverse vitamin B family members, which has extensively antagonistic effects on developmental toxicity and teratogenesis. Lu et al. (
2008) revealed that vitamin B
12 could effectively antagonize DEX-induced failure of the palatal fusion in mice embryos. However, an effect of vitamin B
12 on the combined toxicity of TCDD and DEX for palatal development has not been investigated.
Several growth factors have been reported to influence the complex events during palatogenesis. Among them, transforming growth factor β (TGF-β) has been shown to be critical. Supporting evidence indicated that TGF-β3 and its stimulators hold potential roles as antidotes to TCDD-induced cleft palate (Thomae et al.,
2005). Based on studies using misexpression and chemical inhibition, it has also been suggested that the TGF-β/activin receptor-like kinase 5 (ALK5)/Smad2 pathway has an important function in disintegration of the midline seam (Dudas et al.,
2004; Cui et al.,
2005). ALK5, TGF-β type I receptor, was demonstrated to play an indispensable role in the disappearance of the palatal epithelial seam during palatal fusion, especially in the anterior and posterior aspects of the secondary palate (Dudas et al.,
2006). Based on the reports mentioned above, we believe that TGF-β3 and ALK5 may play an important role in combining teratogenic action of TCDD and DEX.
In the present study, we aimed to identify the protective effect of vitamin B
12 on palatal development deficit induced by co-administration of TCDD and DEX. Here we initially investigated the morphological and histological alterations among TCDD+DEX exposed mice treated with or without vitamin B
12 administration. And then we measured the expression levels of TGF-β3 and ALK5 during palatogenesis.
2. Materials and methods
2.1. Mice breeding and treatment
Adult C57BL/6J mice were from the Laboratory Animal Center of Sichuan University (China) and were bred in a specified-pathogens free (SPF) room, with temperature of 20–25 °C, adequate food, and free drinking water (filtered water). Two adult females were caged overnight with a male and examined in the next morning for the presence of a vaginal plug. The presence of vaginal plugs was designated as Day 0 Hour 0 (GD 0
0) and the females were weighed. On GD 10
0 (or GD 10) these same mice were weighed again and those gaining at least 2 g were assumed pregnant. All experimental procedures were approved and carried out in accordance with the Institutional Animal Care and Use Committee of the Laboratory Animal Center of Sichuan University, China.
The pregnant mice were divided randomly into a mock control (Group A), TCDD+DEX exposed (Group B) and TCDD+DEX+vitamin B
12 exposed group (Group C). The individual GD 12 animal was intraperitoneally injected once a day with 0.1 ml saline (for Group A), 25 mg/kg DEX (for Group B), or 25 mg/kg DEX+10 mg/kg vitamin B
12 (for Group C). At GD 12, TCDD (6 μg/kg) was orally administrated to subjects in both Groups B and C, but saline was use as vehicle control for Group A animals.
The TCDD (Wellington Laboratory, Canada) stock solution was prepared as described by Abbott et al. (
1999). Vitamin B
12 was purchased from Sigma and DEX sodium phosphate was provided by the State Key Laboratory of Oral Diseases, Sichuan University, China.
2.2. Histological analysis and cleft palate assessment
At GD 17.5, six pregnant mice in each group were dissected to calculate the incidence of cleft palate by the naked eye. The embryos at GD 13.5, 14.5, and 15.5 were fixed in 0.04 g/ml paraformaldehyde in phosphate-buffered saline (PBS) at 4 °C overnight. Then the heads of embryos were embedded in paraffin, and 5 μm-thick serial coronal sections were made and stained with hematoxylin and eosin (H&E). Finally, these sections were photographed with a Leica photographic system (Germany). The development of palatal plate was detected quantitatively by measuring the average distance between two cut-off points of the opposing palate shelves in acquired images, including the eye tissue (Fig.
1). In order to make sure of picking the same level of section to compare these three groups, the position of small eye tissue was set up to be a reference of measurement. In the site, the palatal specimen was sequentially cut into fifty sections. Out of every ten sections, one slice was chosen for measurements. Five palate specimens were included in each group.
Fig.1
Measurement of the distance between two cut-off points of opposing palate shelves
Arrowheads indicate eye tissue. N: nasal; P: palate; d: distance. Magnification: 40×
2.3. Scanning electron microscopy (SEM) and palatal surface structure
For observation of the fine surface structure of palatal shelves, we carried out SEM as described by Ding et al. (
2001). Samples were fixed for 2 h in 0.025 g/ml glutaraldehyde solution at room temperature, and subsequently were dehydrated through a graded series of ethanol, isoamylacetate replacement, critical point drying, and gold plating. Lastly, the images were viewed on an SEM (Hitachi S-2460 N, Tokyo, Japan).
2.4. Expression analysis of genes
To test the expression levels of TGF-β3 and ALK5 in embryonic mice palates of different groups, quantitative reverse transcription polymerase chain reaction (RT-PCR) analyses were conducted. Palatal shelves were dissected from staged embryos on GD 13.5, 14.5, and 15.5, and stored at −70 °C. Total RNA was extracted from the excised palatal shelves, RNA was reverse-transcribed to cDNA, and then the resulting cDNA was subjected to a PCR in a Bio-Rad iCycler iQ system. Finally, the comparative cycle threshold (
C
T) was used to analyze the data by generating relative values of the amount of target cDNA as described (Schmittgen and Livak,
2008). The relative amounts of TGF-β3 and ALK5 were determined according to an equation: 2
−∆∆C
T
=2
−∆(C
T, gene of interest−C
T, internal control). In our study, we selected TGF-β3 and ALK5 as target genes and β-actin as internal control.
2.5. Statistical analysis
Data were analyzed using SPSS 13.0 software. The incidence of cleft palate among different groups was compared using the chi-square and Fisher’s exact test. Average distance and quantitative RT-PCR data among groups were analyzed using repeated measures, followed by the least significant difference (LSD) test.
P<0.05 was considered significantly different.
3. Results
3.1. TCDD+DEX expose-induced cleft palate morphology deficit failed to be prevented by vitamin B12
As shown in Table
1, TCDD+DEX exposure induced cleft palate deficit in all embryos (with 100% incidence of cleft palate). Vitamin B
12 failed to reduce the incidence of cleft palate (
P>0.05). Fig.
2 showed that the sizes of the palate among the three groups have no distinguished differences on GD 13.5. Palatal shelves grew vertically downward beside the tongue (Figs.
2a, 2d, and 2g). On GD 14.5 and GD 15.5, the palatal shelves in the control group (Group A) were elevated into a horizontal apposition above the tongue to make contact and fuse. A medial edge epithelial seam was still obvious on GD 14.5 (Figs.
2b and 2c). However, in the treated Groups B and C embryos, the palatal shelves were elevated above the tongue, but failed to meet and fuse and thus displayed the same abnormal growth (Figs.
2e, 2f, 2h and 2i). To explore the antagonistic effect of vitamin B
12 on the combined teratogenicity quantitatively, we calculated the average distance in each group and found that there was no significant difference between Groups B and C, although the distances in Group C were smaller than those in Group B in different periods (Table
2).
Table 1
Fetal mice development and incidence of cleft palate among different groups
| Group |
Pregnant mice |
Live births |
Fetuses with clefts |
Frequency of clefts (%) |
| A |
6 |
36 |
1 |
2.8 |
| B |
6 |
37 |
37 |
100 |
| C |
6 |
34 |
34 |
100 |
Split χ
2 test showed that there was no difference between Groups B and C (P>0.05)
Fig.2
Histological sections of embryonic palate in three groups
(a–c) Group A on GD 13.5, GD 14.5, and GD 15.5, respectively; (d–f) and (g–i) Groups B and C, respectively, at different periods. On GD 13.5, the size and morphology of palate among three groups were similar (a, d, and g). On GD 14.5, the palate shelves in Group A were elevated and already fused. While in Groups B and C, the size was smaller and there was no contact between opposing palate shelves though they could be elevated to normal position (b, e, and h). At the time of GD 15.5, the fused palate was further developed in Group A, but in Groups B and C the two opposing shelves remained separated (c, f, and i). Each picture (100×) was an enlarged view of the frame in the inset (40×). Black arrow indicates fused palate. Scan bar=100 μm
Table 2
Average distance between the two cut-off points of opposing palates among different groups
| Group |
Distance (μm)
|
| GD 13.5 |
GD 14.5 |
GD 15.5 |
| A |
313.3±8.0ac
|
0ad
|
0ad
|
| B |
328.1±10.3ac
|
241.5±7.1bd
|
154.1±47.5be
|
| C |
323.7±9.0ac
|
225.9±12.8bd
|
100.0±13.9be
|
Data are expressed as mean±SD. a–b The same letter denoted values that are not significantly different within the same time period in different groups (P>0.05). c–e The same letter denoted values that are not significantly different at different time periods in the same groups (P>0.05)
3.2. Vitamin B12 partially restored the altered epithelium differentiation
To determine whether vitamin B
12 has any effects on TCDD+DEX-induced epithelium differentiation, we carried out the SEM on palatal epithelium in different groups. On GD 13.5, while there was no obvious difference in palatal sizes among three groups, the surface structure of palatal shelves had already started to change in Groups B and C. In Group A (Fig.
3a), most of the epithelium cells were bulging and had a clear edge among adjacent cells, while in Group B the epithelial cell trans-differentiated, underwent hyperplasia, and formed a stratified, squamous, and shrinking epithelium (Fig.
3d). In Group C, there were a few of the bulging cells on the epithelium, but the size was smaller than that in Group A (Fig.
3g). On GD 14.5, the morphology of epithelium cells in Groups A and C was similar to that on GD 13.5, respectively, while the epithelium cells were partly damaged and desquamated (Fig.
3h). At stage GD 15.5, the fused medial epithelium cells underwent normal gradative epithelial keratinization (Fig.
3c), while in Groups B and C, the epithelium cells showed deposition of a keratin-like layer presenting irregularity; a small rate of cellular desquamation only appeared on Group B (Figs.
3f and 3i).
Fig.3
Medial edge epithelium (MEE) of palatal palates among the three groups at different times using scanning electron microscopy (SEM)
(a–c) Normal cell morphology throughout the palatogenesis in control group embryos. (d–f) The epithelial cell trans-differentiated, underwent hyperplasia, and formed abnormal epithelium in TCDD+DEX exposed embryos. (g–i) There were some differences from that of Group B in the antagonistic group with a few of the bulging cells on the epithelium on GD 13.5 and no obvious damage and desquamated epithelium on GD 14.5 and GD 15.5 (5000×). White arrow indicates fused palate
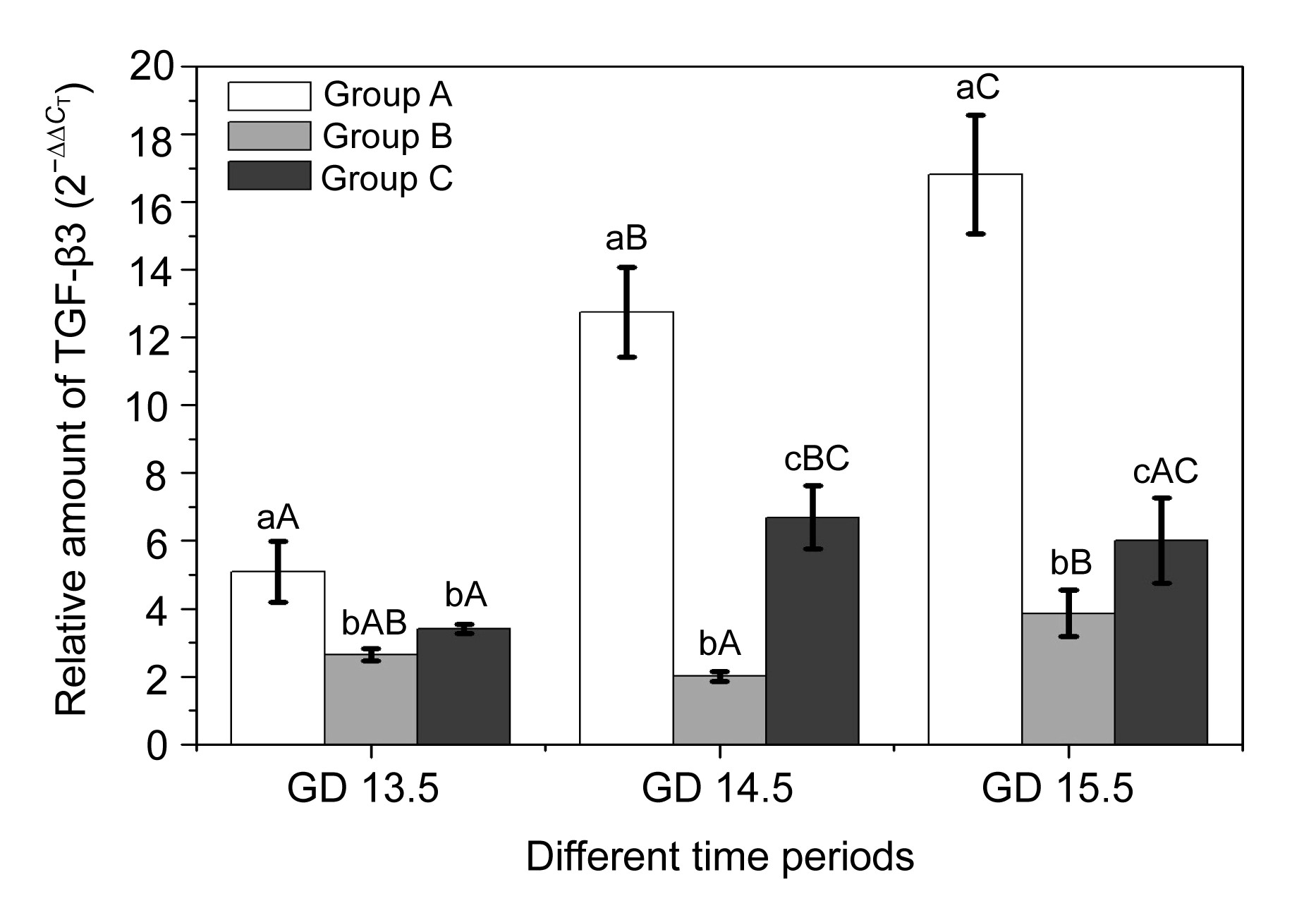
3.3. Vitamin B12 could partly rescue the altered expression levels of TGF-β3 and ALK5
In the present study, we determined the expression levels of TGF-β3 and ALK5 by quantitative RT-PCR. The results showed that the expression of TGF-β3 increased with time, reaching the top on GD 15.5 in Group A, while ALK5 expression decreased to the lowest on GD 15.5. We found that there was a partial correlation between TGF-β3 and ALK5, which showed an opposite expression pattern (Figs.
4 and
5). No similar expression-trends of TGF-β3 and ALK5 were found in Groups B and C. On GD 13.5 there were no significant differences in TGF-β3 and ALK5 expression between these two groups (
P>0.05). On GD 14.5 and GD 15.5, a higher amount of TGF-β3 and a lower amount of ALK5 expression were observed in Group C compared to Group B (
P<0.05). All these results revealed that vitamin B
12 may partially restore the altered expression of TGF-β3.
Fig.4
Time-course of TGF-β3 expression in palatal shelves among three groups
a–c The same letter denoted values that were not significantly different within the same time period in different groups (P>0.05). A–C The same letter denoted values that were not significantly different at different time periods in the same groups (P>0.05)
Fig.5
Time-course of ALK5 expression in palatal shelves among three groups
a–c The same letter denoted values that were not significantly different within the same time period in different groups (P>0.05). A–C The same letter denoted values that were not significantly different at different time periods in the same group (P>0.05)
ALK5 is one of the downstream factors of TGF-β3, and is essential for the TGF-β3 signal pathway. On GD 13.5, ALK5 showed a higher expression level, while with the development of the palate, it exhibited a gradually decreasing expression in all groups.
4. Discussion
As mentioned above, cleft palate incidence gradually increased, though it is not life-threatening. The distorted palatal development seriously affects the quality of life in children including psychological, emotional, and other disorders. Vitamin B
12, one of the most important coenzymes, has been proved to be involved in various biochemical reactions
in vivo (Selhub,
2002). Clinical research suggests that a low level of vitamin B
12 in a pregnant woman’s serum is perhaps related to higher cleft palate embryos (van Rooij et al.,
2003) and epidemiological investigations revealed that pregnant women taking certain doses of vitamin B
12 could have a reduced risk of non-syndromic craniofacial clefts.
In vivo animal experiments showed that the incidence of DEX-induced mouse cleft palate decreased after vitamin B
12 exposure (Natsume et al.,
1986).
Our previous studies also demonstrated that vitamin B
12 effectively restored DEX-induced failure of the palatal fusion in mice embryos (Lu et al.,
2008). In our current study, we found that there was no significant difference in the palatal cleft incidence in Groups B and C (Table
1), and the size of palate between the two groups also had no distinguished difference. It demonstrated that vitamin B
12 failed to rescue TCDD+DEX-induced cleft plate in the present experimental design. However, the results of SEM and signal pathway molecule analysis revealed that vitamin B
12 could protect epithelium cells from damage by combined teratogenic action and restore the altered epithelium differentiation and expressions of TGF-β3 and ALK5 to a certain extent.
As reviewed previously, formation of the mammalian secondary palate is a complex process, including a highly coordinated series of morphogenetic events that are mediated by many more signaling molecules (Zimmerman,
1997). The high incidence of cleft palate in mice induced by TCDD+DEX involves many factors; Abbott (1995) proposed a circulation model to interpret the high cleft palate incidence associated with a few of the growth factors including TGF-β1, epidermal growth factor (EGF), etc. In this study, we found that ALK5 displayed an opposite expression pattern when compared to TGF-β3, which might relate to feedback regulation at the molecular level. A coordinated expression of other genes that regulate cell proliferation, differentiation, transformation, and cell death is required for the development of the palatal palate (Zimmerman,
1997). The current study only defined altered expressions of TGF-β3 and ALK5. It is likely that other factors may be involved in the palatal development as well. Thus, further studies will be needed to elaborate that profile.
In summary, our study suggests that vitamin B
12 may not impact the incidence of cleft palate. However, it affects the expressions of TGF-β3 and ALK5 in TCDD+DEX exposure-induced cleft palate. Along with other existing association studies, our research indicates that there are still many signal molecules participating in TCDD+DEX-induced cleft palate.
* Project supported by the National Natural Science Foundation of China (Nos. 81000425 and 30530730) and the Sichuan Key Technology R&D Program (No. 2010SZ0098), ChinaCompliance with ethics guidelines Shu-fan ZHAO, Mao-zhou CHAI, Min WU, Yong-hong HE, Tian MENG, and Bing SHI declare that they have no conflict of interest.References
[1] Abbott, B.D., Schmid, J.E., Brown, J.G., 1999. RT-PCR quantification of AHR, ARNT, GR, and CYP1A1 mRNA in craniofacial tissues of embryonic mice exposed to 2,3,7,8-tetrachlorodibenzo-
p-dioxin and hydrocortisone.
Toxicol Sci, 47(1):76-85.


[2] Chai, M.Z., Li, C.H., He, Y.H., 2010. The preliminary study on transforming growth factor-β3, activin receptor-like kinase 5 expression in 2,3,7,8-tetrachloro-
p-dibenzodioxin and dexamethasone-induced palatal cleft in mice.
West China J Stomatol, (in Chinese),28(4):356-360.
[3] Cui, X.M., Shiomi, N., Chen, J., 2005. Overexpression of Smad 2 in TGF-β3-null mutant mice rescues cleft palate.
Dev Biol, 278(1):193-202.
[4] Ding, H., Roncari, L., Shannon, P., 2001. Astrocyte-specific expression of activated p21-ras results in malignantastrocytoma formation in a transgenic mouse model of human gliomas.
Cancer Res, 61(9):3826-3836.
[5] Dudas, M., Nagy, A., Laping, N.J., 2004. TGF-β3-induced palatal fusion is mediated by Alk-5/Smad pathway.
Dev Biol, 266(1):96-108.
[6] Dudas, M., Kim, J., Li, W.Y., 2006. Epithelial and ectomesenchymal role of the type I TGF-β receptor ALK5 during facial morphogenesis and palatal fusion.
Dev Biol, 296(2):298-314.
[7] Lu, S.J., He, W., Shi, B., 2008. A preliminary study on the teratogenesis of dexamethasone and the preventive effect of vitamin B
12 on murine embryonic palatal shelf fusion
in vitro
.
J Zhejiang Univ-Sci B, 9(4):306-312.
[8] Murray, J.C., Schutte, B.C., 2004. Cleft palate: players, pathways, and pursuits.
J Clin Invest, 113(12):1676-1678.
[9] Natsume, N., Narukawa, T., Kawai, T., 1986. Teratogenesis of dexamethasone and preventive effect of vitamin B
12
.
Int J Oral Maxillofac Surg, 15(6):752-755.
[10] Schmittgen, T.D., Livak, K.J., 2008. Analyzing real-time PCR data by the comparative
C
T method.
Nat Protoc, 3(6):1101-1108.
[11] Selhub, J., 2002. Folate vitamin B
12 and vitamin B
6 and onecarbon metabolism.
J Nutr Health Aging, 6(1):39-42.
[12] Thomae, T.L., Stevens, E.A., Bradfield, C.A., 2005. Transforming growth factor-β3 restores fusion in palatal shelves exposed to 2,3,7,8-tetrachlorodibenzo-
p-dioxin.
J Biol Chem, 280(13):12742-12746.
[13] van Rooij, I.A., Swinkels, D.W., Blom, H.J., 2003. Vitamin and homocysteine status of mothers and infants and the risk of nonsyndromic orofacial clefts.
Am J Obstet Gynecol, 189(4):1155-1160.
[14] Zimmerman, E.F., 1997. Palate. Handbook of Experimental Pharmacology 124/I: Drug Toxicity in Embryonic Development I. Springer-Verlag,New York :183-208.





Open peer comments: Debate/Discuss/Question/Opinion
<1>