1. Introduction
Gene-promoted bone tissue engineering provides a new treatment strategy for repairing bone defects caused by fractures, bone loss, or tumors (Breitbart et al.,
1999; Mason et al.,
2000; Edwards and Mason,
2006a;
2006b). Three elements are necessary: seed cells for osteogenesis, osteoinductive genes for osteoinduction, and specific scaffolds for osteoconduction. Bone marrow stromal cells (BMSCs) are the most universal source of adult stem cells and play an important role in the field of gene-enhanced tissue engineering in many seed cells (Pittenger et al.,
1999; Lange et al.,
2005; Liu et al.,
2006). Compared with other cells, BMSCs have intrinsic osteogenic properties, but they also have disadvantages: suitable quantities are difficult to obtain
in vitro, and there is a limitation of age-related decline in the number of these cells (Oreffo et al.,
1998; Muschler et al.,
2001; Derubeis and Cancedda,
2004). In contrast, adipose-derived stem cells (ADSCs) are better able to differentiate into adipocytes but less able to differentiate into osteogenic cells. However, ADSCs have received much attention because they constitute an abundant and easily accessible resource of stem cells and have pluripotent lines able to differentiate into adipogenic, osteogenic, myogenic, and chondrogenic cells (Halvorsen et al.,
2001; Zuk et al.,
2001; Nathan et al.,
2003; Fraser et al.,
2006; Schaffler and Buchler,
2007).
Bone regeneration by transplantation of ADSCs must be preceded by
in vitro osteogenic induction to osteoblasts during cell culture by the addition of dexamethasone, bone morphogenic proteins (BMPs), or other factors (Jaiswal et al.,
1997). However, these
in vitro treatments cannot be sustained after implantation
in vivo. Osteogenic differentiation of stem cells with the aid of gene transfection provides a possible solution to these limitations (Asahara et al.,
2000).
Osterix (OSX) is a transcription factor containing a zinc finger encoded by the
Sp7 gene, which is important in osteoblast differentiation and bone generation, activating downstream of
Runx2 to induce preosteoblasts to differentiate into functional osteoblasts (Nakashima et al.,
2002; Lee et al.,
2006; Nishio et al.,
2006). OSX is also a transcription factor essential for osteoblast differentiation because no bone generation occurs in
OSX-null mice (Tai et al.,
2004). Overexpression of
OSX gene has been shown to be sufficient to induce differentiation of ADSCs towards osteoblastic cells
in vitro (Wu et al.,
2007).
As an internal bone tissue engineering technique, distraction osteogenesis (DO) provides an ideal model for the study of bone regeneration. Therefore, we hypothesized that the local delivery of ADSCs transfected with OSX would increase the numbers of osteoblasts in distracted areas and promote osteogenesis in distraction regions.
2. Materials and methods
2.1. Animals
Fifty-four six-month-old New Zealand male rabbits were purchased from the Experimental Animal Center of Shandong University, China. Animals were housed in pathogen-free conditions and all studies performed were in accordance with institutional guidelines at the Animal Care Veterinary Service Facility of Shandong University, China.
2.2. Surgery of distraction osteogenesis
All rabbits received osteotomy and distraction of the right mandibles and were then randomly divided into three groups with 18 rabbits in each group. The rabbit model has been reported in many previous studies dealing with mandibular distraction (Qi et al.,
2006; Hu et al.,
2007; Lai et al.,
2011). Briefly, the rabbits were given penicillin with at a dose of 15 U/kg prior to operation; anesthesia was achieved by intravenous administration of pentobarbital (1 mg/kg) and subcutaneous injection of local anesthesia [1% (w/v) lidocaine]. Surgical procedures were conducted according to Lai et al. (
2011). The distracters (Fig.
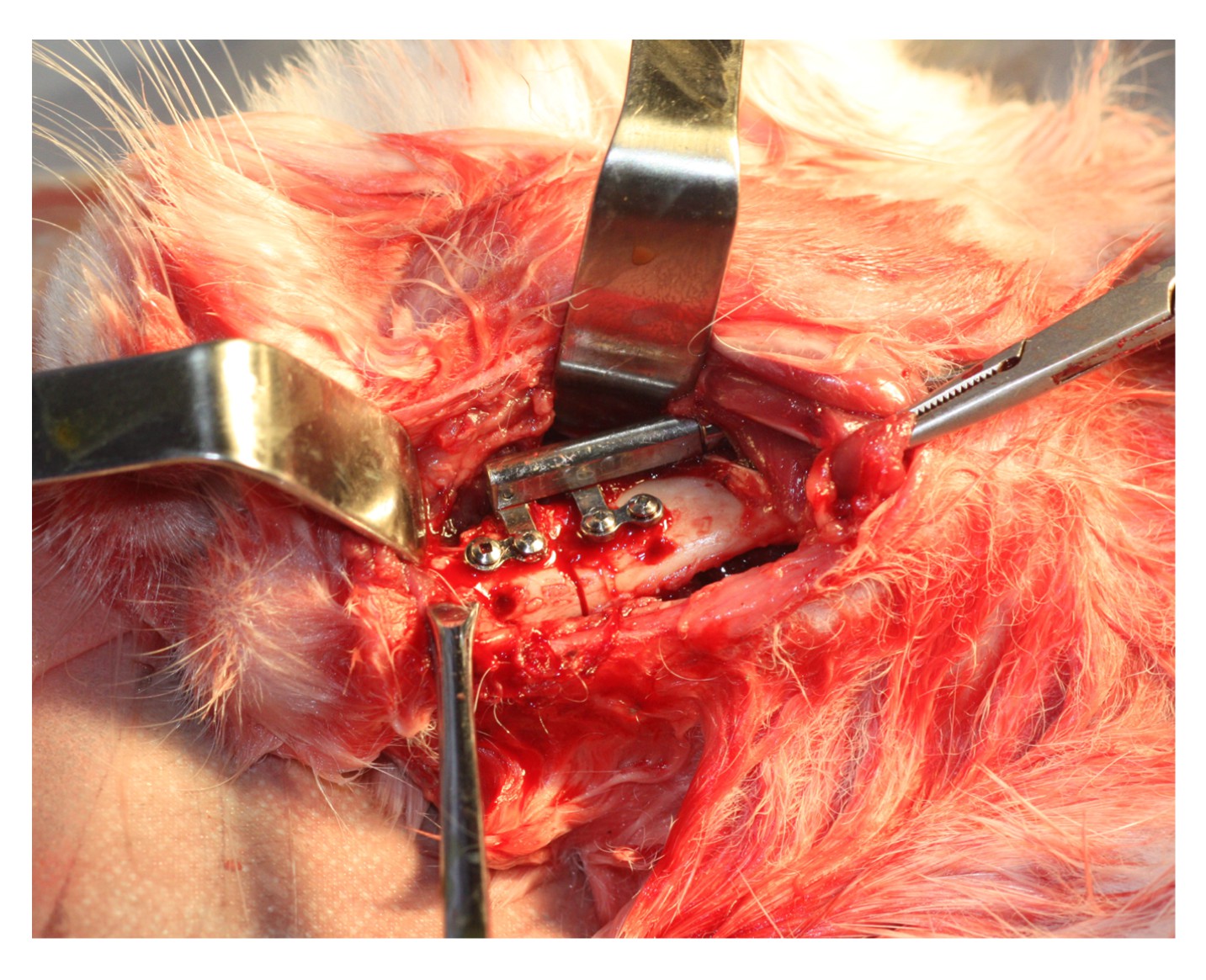
1) were purchased from Xi’an Zhongbang Titanium Biological Materials Co., Ltd. (China). After the distraction surgery, the distracter was fixed in the mandible (Fig.
2). The rabbits were fed on a soft diet for the following three weeks. Their teeth were polished to prevent overgrowth during mandibular distraction.
Fig.1
Custom-made internal distractor purchased from Xi’an Zhongbang Titanium Biological Materials Co., Ltd., China
Fig.2
Custom-made distraction device fixed in situ
2.3. Isolation and culture of ADSCs
A sterile surgical procedure was performed to obtain 10 ml inguinal adipose tissue from the rabbits. To remove contaminating debris, tissues were excised and washed 3–4 times with sterile phosphate-buffered saline (PBS), and then were minced into small pieces and incubated with 0.2% (w/v) type I collagenase (Sigma, USA) in PBS at 37 °C on a shaker for 2 h. After centrifugation of the resultant soup, the pellet with adherent stromal cells was placed in Ficoll solution (Biosera) and centrifuged again. The stromal vascular fraction (SVF) in the second layer was transferred into a tube and washed with PBS. The pellet with SVF was resuspended in culture medium that contained 1% (w/v) penicillin/streptomycin (Biosera) and 10% (w/v) fetal bovine serum (GIBCO). The adherent cells were cultured and non-adherent cells were discarded. The medium was changed every 4 d and cells were harvested after about 30 d of culture.
The recombinant plasmid
pEGFP-OSX (EGFP is the enhanced green fluorescence protein) was constructed by Shanghai Genechem Co., Ltd. (China) using a directed cloning technique. ADSCs were transfected with
pEGFP-OSX using Lipofectamine™ 2000 (Invitrogen, USA) when they grew to 80% confluence, and the transfection efficiency was determined using fluorescence microscopy (Leica, Wetzlar, Germany) on Day 2.
2.4. Therapy
After 6 d of latency, the fractured bones were distracted by 0.8 mm twice a day for 10 d. Immediately after the distraction phase, the distraction areas of the animals in group A received an injection of about 1×10
7
OSX-transfected ADSCs suspended in 0.2 ml physiological saline. Rabbits in group B were injected on the same schedule with 0.2 ml of ADSCs transfected with
pEGFP-N1, and rabbits in group C received an injection of 0.2 ml of physiological saline only. Animals were sacrificed at Week 2 (half of the animals in each group) or at Week 6 (the remaining animals in each group) after surgery.
2.5. Radiographic examination
Lateral radiographs of specimens were taken at two and six weeks after distraction. The radiation dosage (SIEMENS, Germany) was set at 50 kV and 16 mA, with a 0.06-ms exposure time. The distraction bone density was measured as the projectional bone mineral density (BMD).
The distraction gap of each specimen was evenly divided into three parts along the axial plane, and nine evenly distributed points were marked in the distraction gap, three in each part. Their values were standardized according to the mean of two marker points in incisor area of the mandible. The value of each point (
Y) was calculated according to
Y=255×
Z/
X, where
X stands for the mean of the actual brightness at two marker points and
Z stands for the actual brightness of each point. The means of nine points were used as the measure of distraction bone density.
2.6. Histological examination
After the animals were euthanized, gross features of their distraction regions were examined. Specimens were decalcified in a solution of 13% (w/v) ethylenediaminetetraacetic acid (EDTA) buffer (pH 7.2) after fixation. The samples were cut into four pieces along the axial plane longitudinally, and then all pieces were embedded in paraffin. Sections of 5-μm thickness were prepared for hematoxylin and eosin (H&E) staining. Histological analysis was performed in a prospective, randomized, and blinded manner by two experienced pathologists, similar to the methods of Ashinoff et al. (
2004) and Hu et al. (
2007). Three parameters, including the thickness of new trabeculae (TNT; μm), and the volumes of the newly generated cortical bone (NBV1) and the cancellous bone (NBV2) of the distracted regions, were measured in 10 discontinuous slices of each specimen (Ashinoff et al.,
2004; Hu et al.,
2007). The means of TNT, NBV1, and NBV2 for groups A, B, and C were subjected to statistical analysis.
2.7. Statistical analysis
Data are presented as mean±standard deviation (SD). Statistical significance was determined by one-way analysis of variance (ANOVA). Variables were considered significant with
P<0.05. All analyses were performed with SPSS 16.0 for windows.
3. Results
3.1. Gross appearance
All the surgeries were completed successfully and all experimental animals survived without serious complications, such as the non-union of bones and the loss of distracters. Twenty days after consolidation, four rabbits suffered from shin infections, but because the newly reformed bone was strong enough to withstand occlusal force, these animals were not excluded from the experiments. The three groups all exhibited typical changes of unilateral mandibular lengthening, overgrowth of the lower incisors, and obvious crossbite. There were no obvious differences in DO wounds, gross bone volume, or appearance of the distraction zones among groups A, B, and C.
3.2. Transfected cell evaluation
ADSCs transfected with
pEGFP-OSX showed transient expression of EGFP under fluorescence microscopy. The transfection efficiency decreased gradually from about 47% at Day 2, 42% at Day 6, 31% at Day 13, 18% at Day 20, to 13% at Day 27.
3.3. Radiographic results
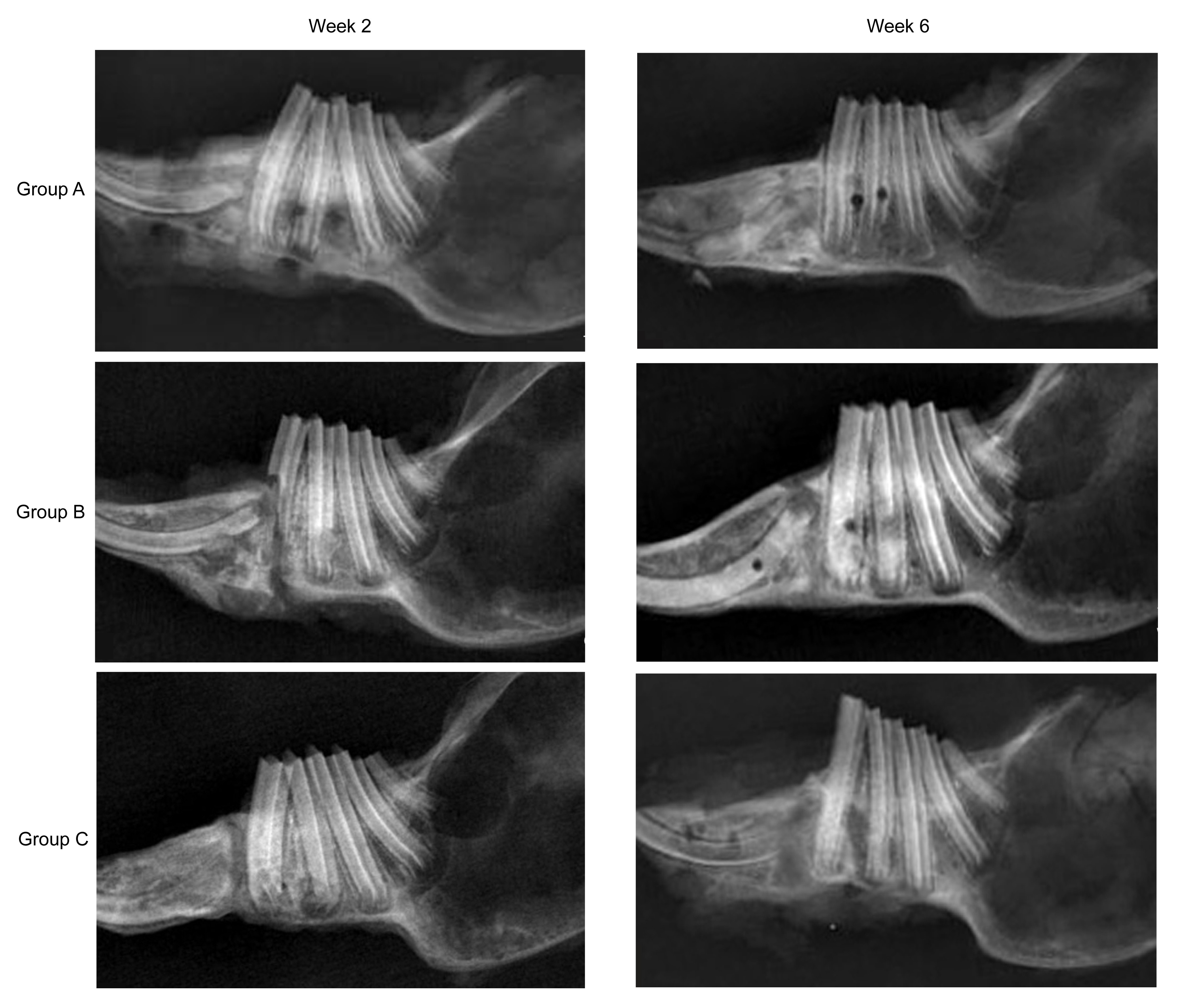
Fig.
3 shows representative lateral radiographs of distracted mandibles at Weeks 2 and 6 after the end of distraction. There were significant differences among groups A, B, and C in terms of gray density analysis. The radiodensity of the distraction areas in group A was higher than that in groups B and C at Weeks 2 and 6 after the distraction procedure.
Fig.3
Lateral radiographs of distracted mandibles in three groups at Weeks 2 and 6 after the end of distraction
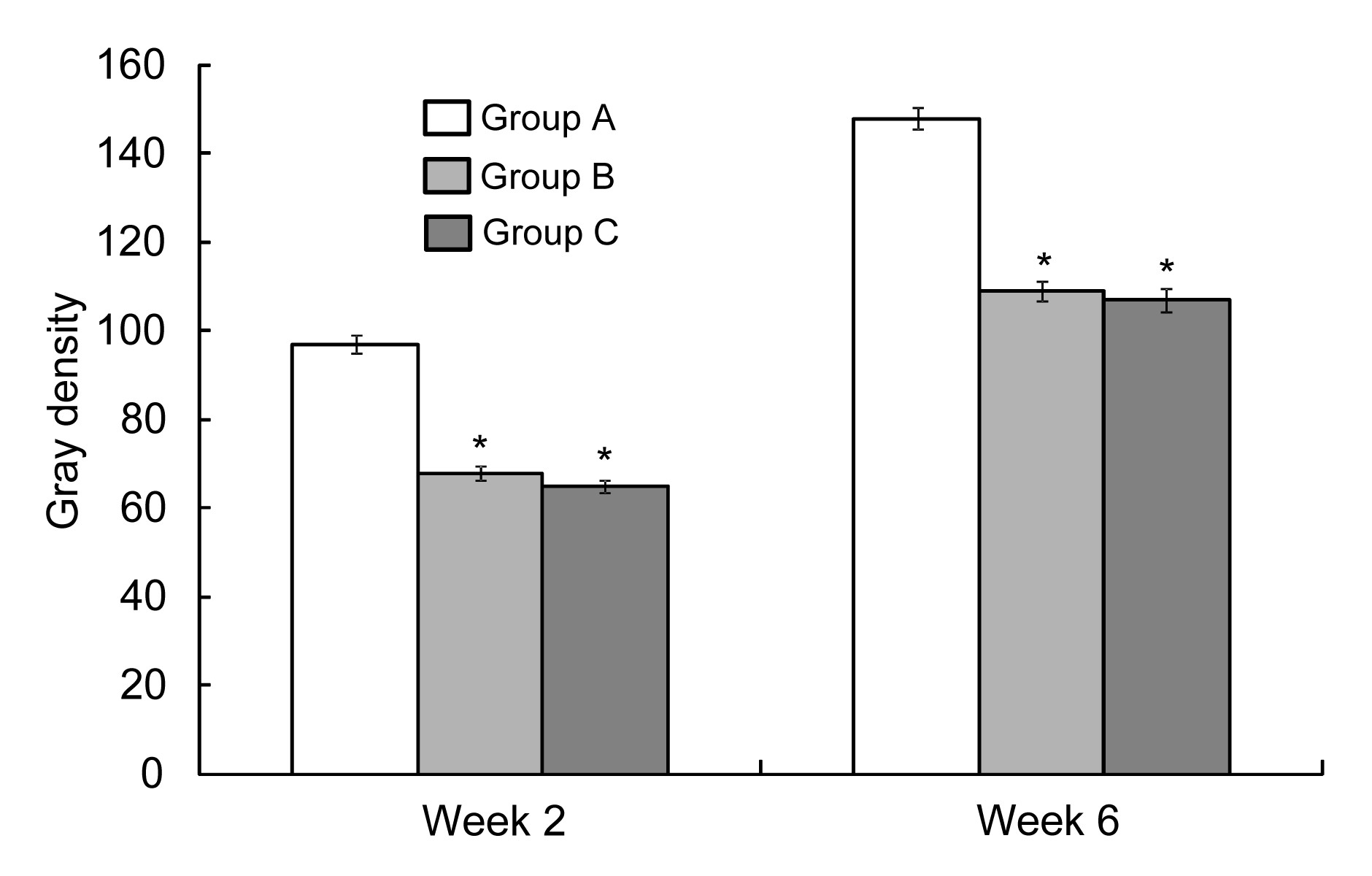
Statistical analysis of the gray density (Fig.
4) also supported these results. The radiodensity of the distracted bone in group A was higher than that in groups B and C (
P<0.01). There was no difference in radiodensity between groups B and C (
P>0.05).
Fig.4
Radiodensity of distracted gaps of different groups at Weeks 2 and 6
*
P<0.01 vs. group A. Data are expressed as mean±SD (n=9)
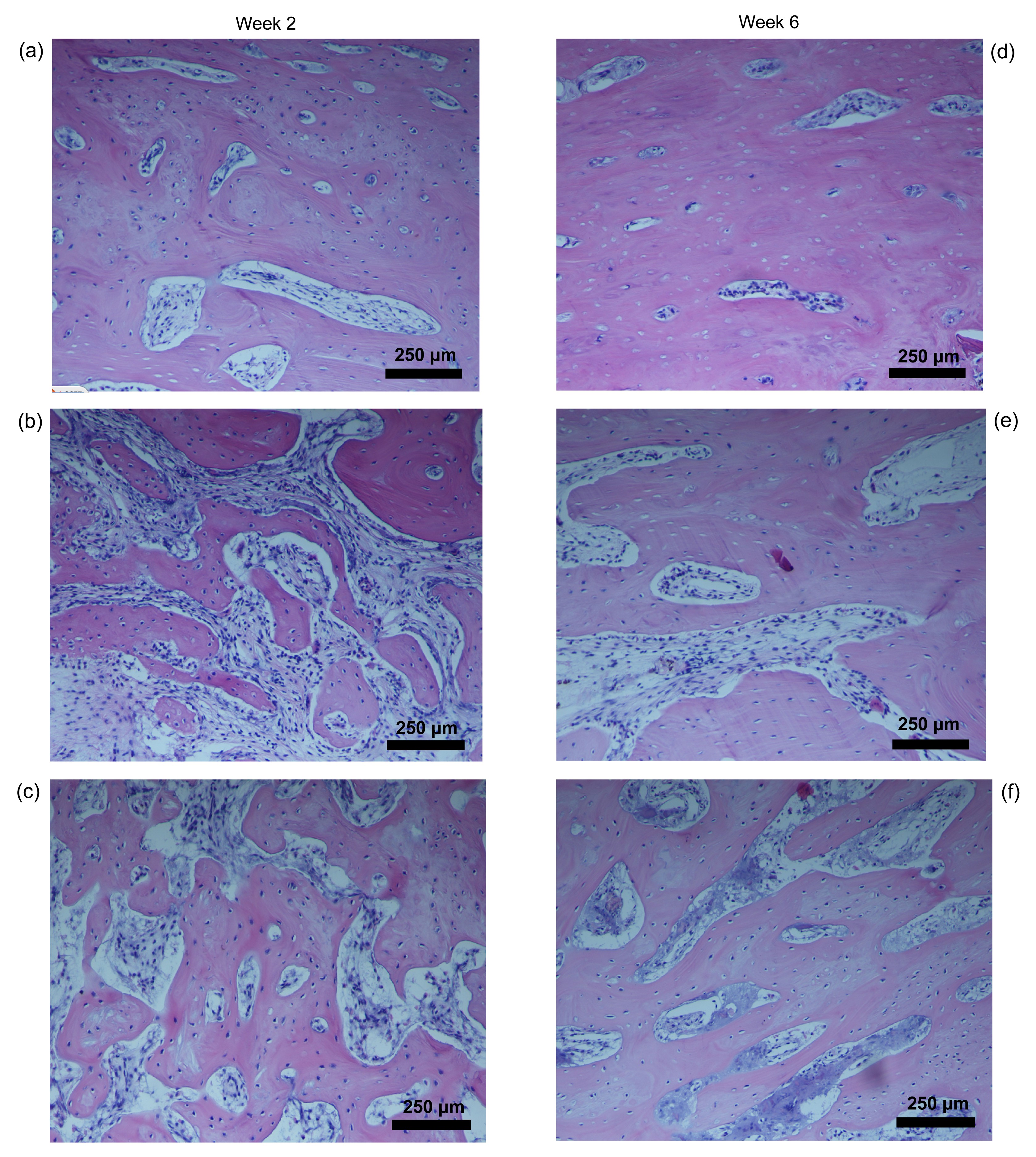
3.4. Histological findings
Light microscopy showed that bone cells in the distracted areas were stretched along in the direction of the distraction. At the second week after the distraction procedure, the two fragments of mandibles in all groups were filled with newly formed bone trabeculae. Similar results were seen in groups B and C, but much denser and thicker bone trabecules were observed in the distracted areas in group A (Fig.
5a) than in group B (Fig.
5b) and group C (Fig.
5c).
Fig.5
Histological behavior of the distracted bone at Weeks 2 and 6 in rabbit mandibles (H&E staining)
(a, d) Group A; (b, e) Groups B; (c, f) Group C
At the sixth week, the distraction gaps of the mandible were full of newly generated bone in all three groups. There was no difference between groups B and C, but group A (Fig.
5d) displayed more and denser bone trabeculae than the other two groups (Figs.
5e and 5f). The values of TNT, NBV1, and NBV2 were significantly greater in group A than in groups B and C (
P<0.05) at both time points (Table
1). There was no significant difference between groups B and C (
P>0.05).
Table 1
Thickness of new trabeculae (TNT) and the volumes of the newly generated cortical bone (NBV1) and the cancellous bone (NBV2) in different groups
| Group |
TNT# (μm)
|
NBV1# (%)
|
NBV2# (%)
|
| Week 2 |
Week 6 |
Week 2 |
Week 6 |
Week 2 |
Week 6 |
| A |
38.79±3.09 |
55.11±2.28 |
62.02±3.04 |
95.92±2.75 |
43.54±2.89 |
67.65±4.14 |
| B |
24.76±3.13*
|
35.61±2.40*
|
45.33±2.54*
|
83.46±2.32*
|
25.65±3.57*
|
41.80±2.81*
|
| C |
23.89±1.86*
|
34.61±3.62*
|
44.83±5.33*
|
83.24±3.16*
|
25.45±3.12*
|
42.08±3.26*
|
|
|
F
|
82.803 |
149.892 |
58.604 |
62.038 |
94.485 |
166.839 |
|
P
|
<0.01 |
<0.01 |
<0.01 |
<0.01 |
<0.01 |
<0.01 |
#Data are expressed as mean±SD (n=9)
*
P<0.01 vs. group A
4. Discussion
The foundation of this study was the successful distraction of fractured bones, which depended on having a suitable animal model. Large-animal models are convenient for distraction surgeries, but lack molecular reagents and the cost is rather high. Although a mouse or rat model may avoid the defects of large animals, such models have other disadvantages: the size of the rat mandible is rather small, the rat has poor tolerance to anesthesia, and there are technical difficulties in using an extremely small distractor compared to those of large-animal models. Our study overcame all such obstacles as that rabbits were suitable for studies of large numbers of animals, and the feeding of rabbits was convenient and at moderate cost. Moreover, there was also sufficient space, in terms of mandibular bone, for surgery.
Previous studies found that BMSCs and ADSCs transfected with
OSX or cultured in osteogenic medium differentiated into functional osteoblasts which changed greatly in shape and size (Gao et al.,
2004; Hu et al.,
2007; Lai et al.,
2011). These osteoblasts originating from ADSCs or BMSCs secreted extracellular matrix and calcified, expressed high levels of alkaline phosphatase (ALP), and eventually formed well-defined mineralized nodules, analogous to ADSCs growing in osteogenic medium. However, the NIH3T3 cell line transfected with
OSX did not show an improved ability for osteoblastic differentiation or mineralization (Kim et al.,
2006), which indicated that osteogenic differentiation was specific to only certain multipotent stem cells. In our study, ADSCs transfected with
pEGFP-OSX in an
in vivo environment promoted osteogenesis greatly during distraction compared with the other two groups. This suggested that ADSCs transfected with
pEGFP-OSX maintained the ability for osteoblastic differentiation in an
in vivo environment. These results also agreed with those of our previous study (Lai et al.,
2011). In that experiment, we used autologous BMSCs as the seed cells. Bone generation
in vivo was achieved by transplanting BMSCs modified by
OSX into the distraction gaps at the end of the distraction, and excellent bone generation in the distraction gaps was observed. This showed that the transplantation of OSX modified BMSCs can promote bone generation in distraction osteogenesis (Lai et al.,
2011).
Adipose tissue provides an abundant source of ADSCs for isolation and use in tissue repair and regeneration (Mason et al.,
2000; Lange et al.,
2005; Liu et al.,
2006; Qi et al.,
2006). ADSCs are easy to maintain and proliferate and their harvesting procedure is less aggressive than that of stem cells derived from bone marrow procedures, giving them a quantitative advantage (Banfi et al.,
2001a;
2001b; Liu et al.,
2012). However, the osteogenic ability of BMSCs is greater than that of ADSCs because BMSCs have intrinsic osteogenic properties, while adipose-derived cells need outside environmental factors to stimulate differentiation into osteogenic cells.
Little is known about the fate of transplanted ADSCs in recipient animals and the factors that induce ADSC migration and differentiation
in vivo. Studies suggested that exposure of ADSCs to the bone microenvironment or to transforming growth factor-β-1 (TGF-β1) before transplantation promoted their survival rates in bone tissues. Also, ADSCs, when exposed to the bone microenvironment, showed high levels of ALP activity. In our study, simply implanting ADSCs into the distraction gaps had no effect, suggesting that unmanipulated ADSCs have a lower ability to migrate and differentiate in bone tissues of the recipient animals. This may be because the microenvironment of the distraction areas did not enhance the survival or differentiation of ADSCs. Thus, pretreatment with TGF-β1 and early exposure of ADSCs to the bone microenvironment to promote cell migration and the differentiation and survival of the cells, are all methods that can be used to enhance osteogenesis.
In the present study, we used scaffold-free allograft ADSCs transfected with
OSX in a rabbit model of DO. Our results showed that the recipient rabbits responded well to the injection of allogeneic ADSCs without immunological reaction. This was possibly due to the anti-inflammatory response and low immunogenicity of ADSCs (Uccelli et al.,
2007).
It has been reported that the immunosuppressive effects of ADSCs are mediated by the secretion of soluble factors such as 3-dioxygenase (IDO), indoleamine 2, interleukin (IL)-10, proataglandins, nitric oxide, hepatocyte growth factor, and TGF-β (Dazzi et al.,
2007). Here, the injection of ADSCs transfected with the
pEGFP-OSX gene has shown clear evidence of promotion of osteogenesis, especially six weeks after the distraction procedure. Statistical analyses of radiological and histological results showed that bone generation was significantly improved during that time. Therefore, their immunosuppressive effects and differentiation potential make ADSCs promising cells for bone tissue repair (Dazzi et al.,
2007).
This
in vivo study indicates that the application of ADSCs transfected with
pEGFP-OSX can improve bone regeneration during DO. Given that it was a short term observation, we believe that studies with larger numbers of other kinds of animal models conducted over longer periods of time can provide more evidence of the effectiveness of this method in future strategies for bone repair and reconstruction. Considering that adipose tissue is easy to harvest and highly abundant, it may become a more promising alternative to bone marrow as a source of stem cells (Zuk et al.,
2002).
5. Conclusions
The results of this study suggest that the transplantation of ADSCs transfected with
pEGFP-OSX can effectively promote bone deposition and callus formation during distraction.
* Project supported by the Seed Fund of the Second Hospital of Shandong University (No. S2013010024), ChinaCompliance with ethics guidelines Qing-guo LAI, Shao-long SUN, Xiao-hong ZHOU, Chen-ping ZHANG, Kui-feng YUAN, Zhong-jun YANG, Sheng-lei LUO, Xiao-peng TANG, and Jiang-bo CI declare that they have no conflict of interest.References
[1] Asahara, T., Kalka, C., Isner, J.M., 2000. Stem cell therapy and gene transfer for regeneration.
Gene Ther, 7(6):451-457.


[2] Ashinoff, R.L., Cetrulo, C.L., Galiano, R.D., 2004. Bone morphogenic protein-2 gene therapy for mandibular distraction osteogenesis.
Ann Plast Surg, 52(6):585-590.
[3] Banfi, A., Bianchi, G., Galotto, M., 2001. Bone marrow stromal damage after chemo/radiotherapy: occurrence, consequences and possibilities of treatment.
Leuk Lymphoma, 42(5):863-870.
[4] Banfi, A., Podest, M., Fazzuoli, L., 2001. High-dose chemotherapy shows a dose-dependent toxicity to bone marrow osteoprogenitors: a mechanism for post-bone marrow transplantation osteopenia.
Cancer, 92(9):2419-2428.
[5] Breitbart, A.S., Grande, D.A., Mason, J.M., 1999. Gene-enhanced tissue engineering: applications for bone healing using cultured periosteal cells transduced retrovirally with the
BMP-7 gene.
Ann Plast Surg, 42(5):488-495.
[6] Dazzi, F., van Laar, J.M., Cope, A., 2007. Cell therapy for autoimmune diseases.
Arthritis Res Ther, 9(2):206
[7] Derubeis, A.R., Cancedda, R., 2004. Bone marrow stromal cells (BMSCs) in bone engineering: limitations and recent advances.
Ann Biomed Eng, 32(1):160-165.
[8] Edwards, P.C., Mason, J.M., 2006. Gene-enhanced tissue engineering for dental hard tissue regeneration: (1) overview and practical considerations.
Head Face Med, 2(1):12
[9] Edwards, P.C., Mason, J.M., 2006. Gene-enhanced tissue engineering for dental hard tissue regeneration: (2) dentin-pulp and periodontal regeneration.
Head Face Med, 2(1):16
[10] Fraser, J.K., Wulur, I., Alfonso, Z., 2006. Fat tissue: an underappreciated source of stem cells for biotechnology.
Trends Biotechnol, 24(4):150-154.
[11] Gao, Y., Jheon, A., Nourkeyhani, H., 2004. Molecular cloning, structure, expression, and chromosomal localization of the human osterix (
SP7) gene.
Gene, 341:101-110.
[12] Halvorsen, Y.D., Franklin, D., Bond, A.L., 2001. Extracellular matrix mineralization and osteoblast gene expression by human adipose tissue-derived stromal cells.
Tissue Eng, 7(6):729-741.
[13] Hu, J., Qi, M.C., Zou, S.J., 2007. Callus formation enhanced by BMP-7
ex vivo gene therapy during distraction osteogenesis in rats.
J Orthop Res, 25(2):241-251.
[14] Jaiswal, N., Haynesworth, S.E., Caplan, A.I., 1997. Osteogenic differentiation of purified, culture-expanded human mesenchymal stem cells
in vitro
.
J Cell Biochem, 64(2):295-312.
[15] Kim, Y.J., Kim, H.N., Park, E.K., 2006. The bone-related Zn finger transcription factor osterix promotes proliferation of mesenchymal cells.
Gene, 366(1):145-151.
[16] Lai, Q.G., Yuan, K.F., Xu, X., 2011. Transcription factor osterix modified bone marrow mesenchymal stem cells enhance callus formation during distraction osteogenesis.
Oral Surg Oral Med Oral Pathol Oral Radiol Endod, 111(4):412-419.
[17] Lange, C., Schroeder, J., Lioznov, M.V., 2005. High-potential human mesenchymal stem cells.
Stem Cells Dev, 14(1):70-80.
[18] Lee, J.Y., Lee, Y.M., Kim, M.J., 2006. Methylation of the mouse
DIx5 and
Osx gene promoters regulates cell type-specific gene expression.
Mol Cells, 22(2):182-188.
[19] Liu, H.Y., Chiou, J.F., Wu, A.T., 2012. The effect of diminished osteogenic signals on reduced osteoporosis recovery in aged mice and the potential therapeutic use of adipose-derived stem cells.
Biomaterials, 33(26):6105-6112.
[20] Liu, W., Cui, L., Cao, Y., 2006. Bone reconstruction with bone marrow stromal cells.
Methods Enzymol, 420:362-380.
[21] Mason, J.M., Breitbart, A.S., Barcia, M., 2000. Cartilage and bone regeneration using gene-enhanced tissue engineering.
Clin Orthop Relat Res, 379:S171-S178.
[22] Muschler, G.F., Nitto, H., Boehm, C.A., 2001. Age- and gender-related changes in the cellularity of human bone marrow and the prevalence of osteoblastic progenitors.
J Orthop Res, 19(1):117-125.
[23] Nakashima, K., Zhou, X., Kunkel, G., 2002. The novel zinc finger-containing transcription factor osterix is required for osteoblast differentiation and bone formation.
Cell, 108(1):17-29.
[24] Nathan, S., De, S.D., Thambyah, A., 2003. Cell-based therapy in the repair of osteochondral defects: a novel use for adipose tissue.
Tissue Eng, 9(4):733-744.
[25] Nishio, Y., Dong, Y., Paris, M., 2006.
Runx2-mediated regulation of the zinc finger
osterix/
Sp7 gene.
Gene, 372(10):62-70.
[26] Oreffo, R.O.C., Bord, S., Triffitt, J.T., 1998. Skeletal progenitor cells and ageing human populations.
Clin Sci, 94(5):549-555.
[27] Pittenger, M.F., Mackay, A.M., Beck, S.C., 1999. Multilineage potential of adult human mesechymal stem cells.
Science, 284(5411):143-147.
[28] Qi, M., Hu, J., Zou, S., 2006. Mandibular distraction osteogenesis enhanced by bone marrow mesenchymal stem cells in rats.
J Craniomaxillofac Surg, 34(5):283-289.
[29] Schaffler, A., Buchler, C., 2007. Concise review: adipose tissue-derived stromal cells-basic and clinical implications for novel cell-based therapies.
Stem Cells, 25(4):818-827.
[30] Tai, G., Polak, J.M., Bishop, A.E., 2004. Differentiation of osteoblasts from murine embryonic stem cells by overexpression of the transcription factor osterix.
Tissue Eng, 10(9-10):1456-1466.
[31] Uccelli, A., Pistoia, V., Moretta, L., 2007. Mesenchymal stem cells: a new strategy for immunosuppression?.
Trends Immunol, 28(5):219-226.
[32] Wu, L., Wu, Y., Lin, Y., 2007. Osteogenic differentiation of adipose derived stem cells promoted by overexpression of osterix.
Mol Cell Biochem, 301(1-2):83-92.
[33] Zuk, P.A., Zhu, M., Mizuno, H., 2001. Multilineage cell from human adipose tissue: implications for cell-based therapies.
Tissue Eng, 7(2):211-228.
[34] Zuk, P.A., Zhu, M., Ashjian, P., 2002. Human adipose tissue is a source of multipotent stem cells.
Mol Biol Cell, 13(12):4279-4295.


Open peer comments: Debate/Discuss/Question/Opinion
<1>