1. Introduction
Sporotrichosis is an acute or chronic mycosis caused by infection of the
Sporothrix schenckii, and recently is demonstrated to be a species complex (López-Romero et al.,
2011). Humans and animals acquire it from inoculation of the fungus through thorns, traumas and scratches or bite wounds (de Lima Barros et al.,
2004; Schubach et al.,
2008), even the spores can enter through the respiratory tract. Sporotrichosis may develop as a disseminated disease in immunocompromised individuals particularly in acquired immunodeficiency syndrome (AIDS) patients (Yap,
2011). Sporotrichosis has a global distribution. In China, it is more frequently encountered in the northeast provinces including Jilin, Heilongjiang, and Liaoning (Zhang and Lin,
2008; Mei et al.,
2011; Song et al.,
2011a;
2011b).
S. schenckii had long been believed to be the unique causal agent of sporotrichosis. Identifications were made primarily based on morphological characters (Lopes-Bezerra et al.,
2006). Phylogenetic analysis based on the sequences of the internal transcribed spacer (ITS) region suggested that more than one species existed in this morphologically defined group, indicating this is a species complex (de Beer et al.,
2003). Recently, isolates received as
S. schenckii were regrouped into at least six cryptic species by multilocus phylogenetic analysis, and exhibited a degree of geographical specificity (Marimon et al.,
2006;
2007; Madrid et al.,
2010). Different clinical forms of sporotrichosis are related to the host condition, the route of infection, and the virulence of pathogen, while the virulence was suggested to be associated with different species of
Sporothrix (Arrillaga-Moncrieff et al.,
2009; Fernandes et al.,
2013). Therefore, it is important to recognize what are the real causal agents of the sporotrichosis in various regions, because of the significant differences in the guiding clinical management among these species. Clinical
Sporothrix species have previously been classified based on the partial calmodulin sequences (Marimon et al.,
2007;
2008b; Tan et al.,
2013), but calmodulin regions have been reported to have a lower amplification efficiency as compared to more commonly used regions such as the ribosomal ITS and β-tubulin. ITS is widely used in fungal identification, and recommended as the general barcode for fungi, although additional markers might be needed to distinguish species in certain complexes (Schoch et al.,
2012). Zhou et al. (
2013) concluded that all the
Sporothrix complex species of clinical isolates could readily be recognized by ITS phylogeny.
The aims of this study were to (1) collect a wide range representative strains of pathogens causing sporotrichosis in China, and (2) characterize these strains and confirm their identity through analyses of the ITS sequences and β-tubulin of these strains with relevant taxa.
2. Materials and methods
2.1. Fungal isolates
Sixty-four clinical isolates identified as
S. schenckii sensu lato were included in this study (Table
1). All these isolates were collected from the Dermatology Departments of three hospitals in China, mostly from farmers, housewives, or children. The breakdown is as follows: 26 from the First Hospital of Jilin University (Northeast China), 13 from the First Hospital of Beijing University (North China), and 25 from the Southwest Hospital of the Third Military Medical University (Southwest China). These isolates were subcultured on 2% potato-dextrose agar plates at 25 °C for 14 d. Isolates were then stored in sterilized water at 4 °C and on potato dextrose agar (PDA) slant at room temperature.
Table 1
Fungal species, strain numbers, origins, GenBank accession numbers, and references used in this study
| Fungal species |
Strain No. |
Origin |
GenBank No.
|
Reference |
| ITS |
β-tubulin |
|
S. globosa
|
LC2401 |
Clinical, Jilin, China |
JX997723 |
KC407844 |
This study |
|
S. globosa
|
LC2402 |
Clinical, Jilin, China |
JX997718 |
KC407845 |
This study |
|
S. globosa
|
LC2403 |
Clinical, Jilin, China |
JX997741 |
KC407846 |
This study |
|
S. globosa
|
LC2404 |
Clinical, Jilin, China |
JX997737 |
KC407847 |
This study |
|
S. globosa
|
LC2405 |
Clinical, Jilin, China |
JX997702 |
KC407848 |
This study |
|
S. globosa
|
LC2406 |
Clinical, Jilin, China |
JX997734 |
KC407911 |
This study |
|
S. globosa
|
LC2407 |
Clinical, Jilin, China |
JX997738 |
KC407849 |
This study |
|
S. globosa
|
LC2408 |
Clinical, Jilin, China |
JX997715 |
KC407850 |
This study |
|
S. globosa
|
LC2409 |
Clinical, Jilin, China |
JX997740 |
KC407851 |
This study |
|
S. globosa
|
LC2410 |
Clinical, Jilin, China |
JX997711 |
KC407852 |
This study |
|
S. globosa
|
LC2411 |
Clinical, Jilin, China |
JX997731 |
KC407853 |
This study |
|
S. globosa
|
LC2412 |
Clinical, Jilin, China |
JX997733 |
KC407854 |
This study |
|
S. globosa
|
LC2413 |
Clinical, Jilin, China |
JX997713 |
KC407855 |
This study |
|
S. globosa
|
LC2414 |
Clinical, Jilin, China |
JX997712 |
KC407856 |
This study |
|
S. globosa
|
LC2415 |
Clinical, Jilin, China |
JX997730 |
KC407857 |
This study |
|
S. globosa
|
LC2416 |
Clinical, Jilin, China |
JX997709 |
KC407858 |
This study |
|
S. globosa
|
LC2417 |
Clinical, Jilin, China |
JX997710 |
KC407859 |
This study |
|
S. globosa
|
LC2418 |
Clinical, Jilin, China |
JX997724 |
KC407860 |
This study |
|
S. globosa
|
LC2419 |
Clinical, Jilin, China |
JX997719 |
KC407861 |
This study |
|
S. globosa
|
LC2420 |
Clinical, Jilin, China |
JX997720 |
KC407862 |
This study |
|
S. globosa
|
LC2421 |
Clinical, Jilin, China |
JX997728 |
KC407863 |
This study |
|
S. globosa
|
LC2422 |
Clinical, Jilin, China |
JX997698 |
KC407864 |
This study |
|
S. globosa
|
LC2423 |
Clinical, Jilin, China |
JX997739 |
KC407865 |
This study |
|
S. globosa
|
LC2424 |
Clinical, Jilin, China |
JX997729 |
KC407866 |
This study |
|
S. globosa
|
LC2425 |
Clinical, Jilin, China |
JX997735 |
KC407867 |
This study |
|
S. globosa
|
LC2426 |
Clinical, Jilin, China |
JX997700 |
KC407868 |
This study |
|
S. globosa
|
LC2427 |
Clinical, Chongqing, China |
JX997722 |
KC407869 |
This study |
|
S. globosa
|
LC2428 |
Clinical, Chongqing, China |
JX997692 |
KC407870 |
This study |
|
S. globosa
|
LC2429 |
Clinical, Chongqing, China |
JX997725 |
KC407871 |
This study |
|
S. globosa
|
LC2430 |
Clinical, Chongqing, China |
JX997717 |
KC407872 |
This study |
|
S. globosa
|
LC2431 |
Clinical, Chongqing, China |
JX997732 |
KC407873 |
This study |
|
S. globosa
|
LC2432 |
Clinical, Chongqing, China |
JX997704 |
KC407874 |
This study |
|
S. globosa
|
LC2433 |
Clinical, Chongqing, China |
JX997701 |
KC407875 |
This study |
|
S. globosa
|
LC2434 |
Clinical, Chongqing, China |
JX997705 |
KC407876 |
This study |
|
S. globosa
|
LC2435 |
Clinical, Chongqing, China |
JX997707 |
KC407877 |
This study |
|
S. globosa
|
LC2436 |
Clinical, Chongqing, China |
JX997742 |
KC407878 |
This study |
|
S. globosa
|
LC2437 |
Clinical, Chongqing, China |
JX997708 |
KC407879 |
This study |
|
S. globosa
|
LC2438 |
Clinical, Chongqing, China |
JX997721 |
KC407880 |
This study |
|
S. globosa
|
LC2439 |
Clinical, Chongqing, China |
JX997703 |
KC407881 |
This study |
|
S. globosa
|
LC2440 |
Clinical, Chongqing, China |
JX997695 |
KC407882 |
This study |
|
S. globosa
|
LC2442 |
Clinical, Chongqing, China |
JX997697 |
KC407884 |
This study |
|
S. globosa
|
LC2443 |
Clinical, Chongqing, China |
JX997693 |
KC407885 |
This study |
|
S. globosa
|
LC2444 |
Clinical, Chongqing, China |
JX997694 |
KC407886 |
This study |
|
S. globosa
|
LC2445 |
Clinical, Chongqing, China |
JX997726 |
KC407887 |
This study |
|
S. globosa
|
LC2446 |
Clinical, Chongqing, China |
JX997727 |
KC407888 |
This study |
|
S. globosa
|
LC2447 |
Clinical, Chongqing, China |
JX997716 |
KC407889 |
This study |
|
S. globosa
|
LC2448 |
Clinical, Chongqing, China |
JX997696 |
KC407890 |
This study |
|
S. globosa
|
LC2449 |
Clinical, Chongqing, China |
JX997699 |
KC407891 |
This study |
|
S. globosa
|
LC2450 |
Clinical, Chongqing, China |
JX997714 |
KC407892 |
This study |
|
S. globosa
|
LC2451 |
Clinical, Chongqing, China |
JX997736 |
KC407893 |
This study |
|
S. globosa
|
LC2452 |
Clinical, Chongqing, China |
JX997706 |
KC407894 |
This study |
|
S. globosa
|
LC2453 |
Clinical, Beijing, China |
KC407830 |
KC407895 |
This study |
|
S. globosa
|
LC2454 |
Clinical, Beijing, China |
KC407831 |
KC407896 |
This study |
|
S. globosa
|
LC2455 |
Clinical, Beijing, China |
KC407832 |
KC407897 |
This study |
|
S. globosa
|
LC2456 |
Clinical, Beijing, China |
KC407833 |
KC407898 |
This study |
|
S. globosa
|
LC2459 |
Clinical, Beijing, China |
KC407834 |
KC407900 |
This study |
|
S. globosa
|
LC2460 |
Clinical, Beijing, China |
KC407835 |
KC407901 |
This study |
|
S. globosa
|
LC2462 |
Clinical, Beijing, China |
KC407837 |
KC407903 |
This study |
|
S. globosa
|
LC2463 |
Clinical, Beijing, China |
KC407838 |
KC407904 |
This study |
|
S. globosa
|
LC2465 |
Clinical, Beijing, China |
KC407839 |
KC407906 |
This study |
|
S. globosa
|
LC2466 |
Clinical, Beijing, China |
KC407840 |
KC407907 |
This study |
|
S. globosa
|
LC2467 |
Clinical, Beijing, China |
KC407841 |
KC407908 |
This study |
|
S. globosa
|
LC2468 |
Clinical, Beijing, China |
KC407842 |
KC407909 |
This study |
|
S. globosa
|
LC2469 |
Clinical, Beijing, China |
KC407843 |
KC407910 |
This study |
|
S. schenckii
|
CBS 117842 (CMW 7614) |
Clinical, South Africa |
AY280495 |
AY280477 |
Aghayeva et al. (2004) |
|
S. schenckii
|
CMW 7615 |
Clinical, South Africa |
AY280496 |
AY280478 |
Aghayeva et al. (2004) |
|
S. schenckii
|
CMW 7612 |
Clinical, South Africa |
AY280494 |
AY280476 |
Aghayeva et al. (2004) |
|
S. schenckii
|
ATCC 26331 (CBS 359.36*
) |
Clinical, USA |
FJ545232 |
AM116911 |
Marimon et
al. (2006) |
|
S. schenckii
|
CMW 5681 |
Clinical, South Africa |
EF127886 |
EF139107 |
de Meyer et al. (2008) |
|
S. brasiliensis
|
IPEC 17943 |
Clinical, Brazil |
FN549902 |
AM116935 |
Madrid et al. (2010) |
|
S. brasiliensis
|
IPEC 15572 |
Clinical, Brazil |
FN549904 |
AM116955 |
Madrid et al. (2010) |
|
S. globosa
|
FMR 8597 |
Clinical, Spain |
FN549904 |
AM116964 |
Madrid et al. (2010) |
|
S. globosa
|
CBS 120340* (FMR 8600)
|
Clinical, Spain |
FN549905 |
AM116966 |
Madrid et al. (2010) |
|
S. luriei
|
KMU 2787 (CBS 937.72*) |
Clinical, South Africa |
AB128012 |
AM747289 |
Kawasaki et al. (2003) |
|
S. mexicana
|
CBS 120341*
|
Environmental, Mexico |
FN549906 |
AM498344 |
Madrid et al. (2010) |
|
S. pallida
|
CBS 150.87* (CMW 17168)
|
Environmental, Germany |
EF127879 |
EF139109 |
de Meyer et al. (2008) |
|
Ophiostoma nigrocarpum
|
CMW 650 |
Abies sp. |
AY280489 |
AY280479 |
Aghayeva et al. (2004) |
*Indicates the type strain
CBS: culture collection of the Centraalbureau voor Schimmelcultures, Utrecht, the Netherlands; CMW: culture collection of the Forestry and Agricultural Biotechnology Institute (FABI), University of Pretoria, Pretoria, South Africa; ATCC: American Type Culture Collection, Virginia, USA; IPEC: Instituto de Pesquisa Clínica Evandro Chagas, Fiocruz, Brazil; FMR: Facultate de Medicina i Ciències de la Sault, Reus, Spain; KMU: Kanazawa Medical University, Ishikawa, Japan
2.2. DNA extraction, amplification, and sequencing
Sporothrix spp. strains were inoculated on 2% PDA plates and incubated at 25 °C for 10 d. About 200 mg of fresh filamentous mycelia were scraped off for DNA extraction. The total genomic DNA was extracted using the cetyltrimethyl ammonium bromide (CTAB) method (Porebski et al.,
1997). The quality and quantity of genomic DNA were detected by using 1% agarose gel electrophoresis imaging. Products were stored at −20 °C. The
BT2a/
BT2b primers were used to amplify the partial β-tubulin gene (Glass and Donaldson,
1995). The ITS region was amplified using the primer pair
ITS1/
ITS4 (White et al.,
1990). The polymerase chain reaction (PCR) mixture consisted of 2.5 μl of 10 mmol/L PCR buffer, 2 μl of 2.5 mmol/L dNTP mix, 1 μl of each primer (10 μmol/L), 1 μl of DNA template, 0.2 μl Taq DNA polymerase, and 17.3 μl double-distilled water. Amplification was performed through the following steps: initial denaturation at 95 °C for 5 min, followed by 35 cycling consisting of denaturing at 95 °C for 30 s, annealing at 52 °C (
Bt2) or 54 °C (ITS) for 30 s, and extension at 72 °C for 1 min, followed by 10 min at 72 °C for extension. The PCR products were quantified by 1% agarose gel electrophoresis, and purified products were sequenced at the Beijing Genomics Institute (BGI; China) with the
ITS1/
ITS4 and
BT2a/
BT2b primers mentioned above.
2.3. Sequence alignment and phylogenetic analysis
Sequences generated from the forward and reverse primers were assembled using a Contig Express component to obtain consensus sequence. The sequence identified for all our isolates was verified by a BLAST search (
http://blast.ncbi.nlm.nih.gov). Several ITS regions and β-tubulin sequences published in GenBank were retrieved as reference sequences, together with sequences of new isolates (Table
1).
Alignments of ITS and combined ITS and β-tubulin were made using an MAFFT program (
http://www.ebi.ac.uk/Tools/msa/mafft/) (Katoh and Toh,
2010), and followed by manual adjustments in BioEdit V7.0.5 (Hall,
1999). Both alignments were used for phylogenetic analyses.
The maximum parsimony (MP) analysis was performed using PAUP V.4.0b10 (Swofford,
2003). Ambiguously aligned regions were excluded from all analysis. Trees were inferred using the heuristic search by the 1000 random sequence addition and the tree-bisection-reconnection (TBR) branch-swapping algorithm. Branches of zero length were collapsed and all minimal-length trees were saved. Gaps were treated as missing data. Clade stability was assessed in a bootstrap test with 1000 replications. Tree length (TL), consistency index (CI), retention index (RI), rescaled consistency index (RC), and homoplasy index (HI) were calculated for the generated parsimony trees. A Kishino-Hasegawa (KH) test was performed to determine whether the trees were significantly different. The trees were visualized in TreeView (Page,
1996). The model of evolution was estimated by using MrModeltest 2.3 (Nylander,
2004).
The newly generated sequences in this study were deposited in the GenBank, with their accession numbers listed in Table
1.
3. Results
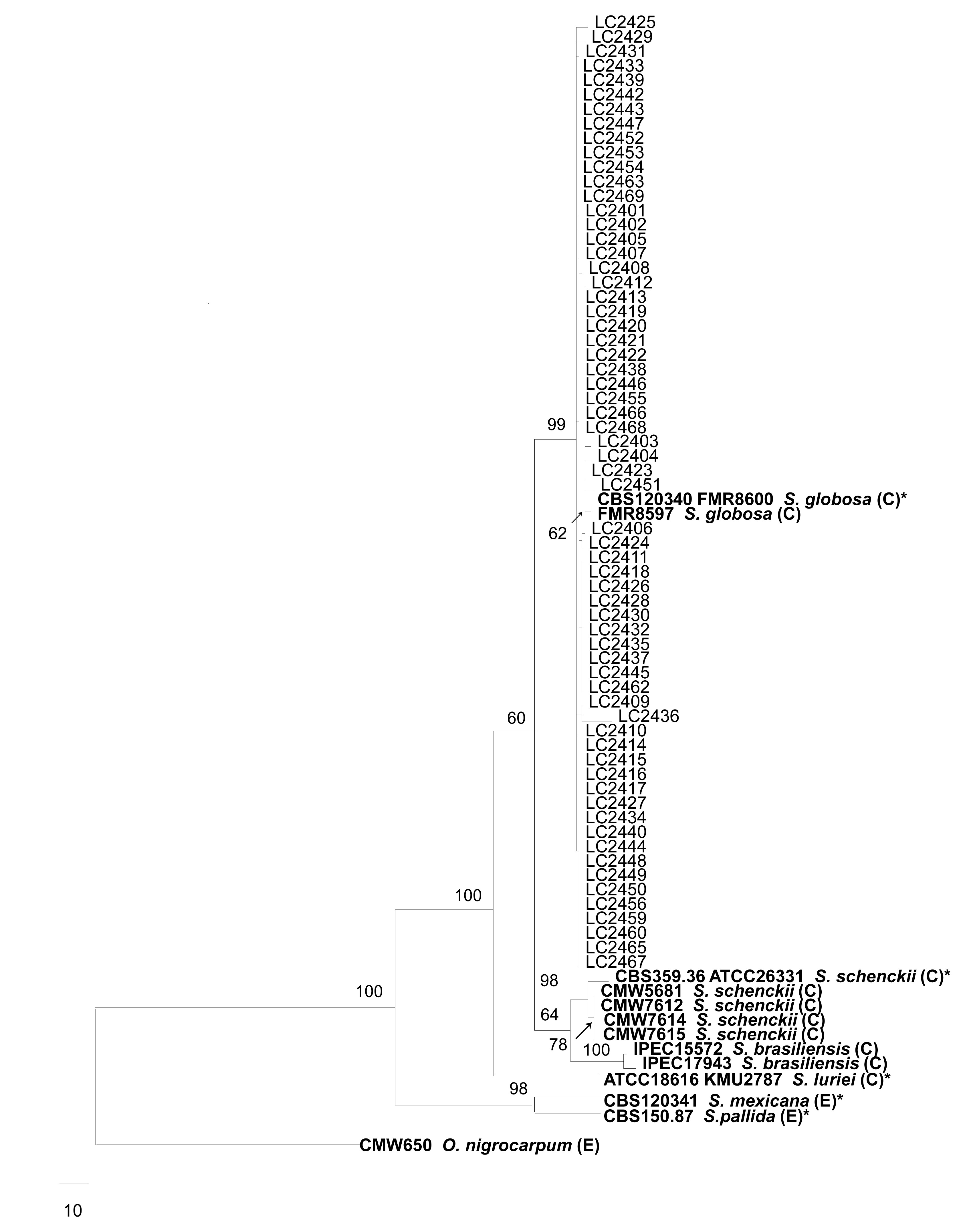
Full length ITS regions and partial β-tubulin genes were amplified in this study. The amplified DNA fragments of the ITS regions and β-tubulin genes were approximatively 450 and 600 bp, respectively. The combined dataset of the ITS regions and β-tubulin genes contained 1052 characters, of which 742 characters were constant, 191 variable characters were parsimony-uninformative, and 119 were parsimony-informative. One of the most parsimonious trees with the shortest tree length (TL=465, CI=0.856, RI=0.835, RC=0.714, and HI=0.144) is shown in Fig.
1. In our combined phylogenetic tree, all the 64 clinical strains collected from different regions of China clustered in a well supported (99%) phylogenetic clade together with the ex-type strain of
S. globosa (CBS 120340). This clade shows sufficient distance to the type of
S. schenckii (CBS 359.36) (Fig.
1). The 64 isolates from China in the
S. globosa clade were distributed among 18 different haplotypes.
Fig.1
Phylogenetic tree generated from a maximum parsimony analysis based on the combined ITS and partial β-tubulin sequence alignment
Bootstrap support values above 50% are shown above the branches, and the scale bar indicates 10 changes. The tree is rooted with Ophiostoma nigrocarpum. Reference sequences are shown in bold. “C” and “E” indicate clinical and environmental strains, respectively. * Indicates the type strain
4. Discussion
S. schenckii was attributed to a unique species as the pathogen of sporotrichosis since it was first isolated from a patient (Lopes-Bezerra et al.,
2006). In past years, various molecular approaches have demonstrated that isolates of
S. schenckii which were identified through morphology displayed diverse genetic characteristics and phylogenetic groupings (Lin et al.,
2000; Mesa-Arango et al.,
2002). Genotyping by analysis of restriction fragment length polymorphism (RFLP) of mitochondrial DNA (mtDNA) and ribosomal DNA (rDNA) demonstrated the existence of intraspecific genetic variability, and all of the four rDNA types showed different geographic distributions (Watanabe et al.,
2004). With molecular phylogenetic analysis,
S. schenckii was discovered to represent more than one species (de Beer et al.,
2003). Marimon et al. (
2007) revealed that
S. schenckii was a complex of at least six phylogenetic species, comprising currently four clinical relevant species, i.e.,
S. schenckii s. str.,
S. brasiliensis,
S. globosa (Marimon et al.,
2007), and
S. luriei (Marimon et al.,
2008b). Marimon et al. (
2007) and Zhou et al. (
2013) both revealed the existence of the correlation between geographical distribution and genotypes among species of
Sporothrix.
S. brasiliensis was confirmed to be restricted to Brazil, while
S. schenckii was mainly prevalent in the Americas, Africa, and Asia (Marimon et al.,
2007; Zhou et al.,
2013).
S. globosa had been reported with widespread geographical distributions, such as in Chile, India, Italy, Japan, Spain, UK, and USA (Madrid et al.,
2009). Previously, based on calmodulin (CAL) sequence analysis, five Chinese environmental isolates received as
S. schenckii had been clustered into the clade of
S. globosa (Marimon et al.,
2007), together with six clinical isolates of
Sporothrix using ITS region analysis (Zhou et al.,
2013), and a total of 11 (100%) Chinese isolates were confirmed to be
S. globosa. In this study, we sampled 64 clinical strains from a wider geographic regions of China, and our combined analysis of ITS and β-tubulin regions further confirmed that the causal agent of sporotrichosis is
S. globosa in China. However, none of these isolates exhibited the same haplotype as the type of
S. globosa CBS 120340, which is a clinical strain from Spain. We did not find a significant correlation between geographic origins of these strains and their genetic variations based on the haplotype data. More research is needed to indicate whether
S. schenckii or other
Sporothrix species exist in China.
Sporotrichosis is characterized by a wide range of cutaneous and extracutaneous clinical manifestations. Kong et al. (
2006) revealed that different randomly amplified polymorphic DNA (RAPD) genotypes of strains led to different clinical forms. There have been reports about cutaneous dissemination and invasive sporotrichosis caused by
S. schenckii and
S. brasiliensis since Marimon et al. (
2007) discovered the three new species of
Sporothrix (Galhardo et al.,
2010; Silva-Vergara et al.,
2012). However, no case of such clinical classification attributed to
S. globosa has been reported. Except of the absence of sporotrichosis forms caused by
S. globosa in some studies (Zhou et al.,
2013), all of the clinical patterns in our study were characterized as fixed and lymphocutaneous, as well as five cases caused by
S. globosa described in Brazil (de Oliveira et al.,
2010; Rodrigues et al.,
2012) and Chile (Cruz et al.,
2012). We considered that this observation could be explained by the different virulence levels of members in
S. schenckii complex. Although a study on the murine model has shown that the most virulent species was
S. brasiliensis, followed by
S. schenckii and
S. globosa (Arrillaga-Moncrieff et al.,
2009), further studies are needed to investigate the possible pathogenesis of different clinical types of sporotrichosis.
Some
in vitro studies of
S. schenckii sensu lato showed variable antifungal susceptibility among different isolates (Brandsberg and French,
1972; McGinnis et al.,
2001), which indicates the existence of cryptic species with variations in antifungal susceptibility. Similar conclusions were presented by the studies of other fungal complexes such as the
Pseudallescheria boydii (Gilgado et al.,
2006) and the
Coccidioides immitis complex (Koufopanou et al.,
2001). In a recent
in vitro antifungal susceptibility test,
S. brasiliensis showed the best response to antifungals in general, followed by
S. schenckii,
S. albicans,
S. globosa, and
S. mexicana (Marimon et al.,
2008a). Therefore, species identification of clinical isolates is essential to determine the effective treatment, and it is significant to study their epidemiology, geographic distribution, and the relevance among taxa, virulence, and clinical patterns.
5. Conclusions
The data in our study showed that the sporotrichosis cases in China were caused by
S. globosa rather than
S. schenckii, and it is demonstrated that ITS and β-tubulin are also efficient to distinguish species in
S. schenckii species complex. With the increasing multilocus sequence data in this group, efficient molecular diagnosis protocols using specific primers should be developed to enable a rapid and accurate identification of clinical
Sporothrix species.
Acknowledgements
The authors acknowledge Ms. Fang LIU (State Key Laboratory of Mycology, Chinese Academy of Sciences) for her assistance in sequencing and data analyses, and Mr. Lei CAI (State Key Laboratory of Mycology, Chinese Academy of Sciences) for his critical comments on this manuscript. We also thank the Dermatology Department of the First Hospital of Jilin University, the Beijing University First Hospital, and the Southwest Hospital of the Third Military Medical University for supplying the clinical strains used in this study.
* Project supported by the National Natural Science Foundation of China (No. 31270062) and the Chongqing Science and Technology Commission (No. cstc2011jjA 10089), ChinaCompliance with ethics guidelines Ting-ting LIU, Ke ZHANG, and Xun ZHOU declare that they have no conflict of interest.References
[1] Aghayeva, D.N., Wingfield, M.J., de Beer, Z.W., 2004. Two new
Ophiostoma species with
Sporothrix anamorphs from Austria and Azerbaijan.
Mycologia, 96(4):866-878.


[2] Arrillaga-Moncrieff, I., Capilla, J., Mayayo, E., 2009. Different virulence levels of the species of
Sporothrix in a murine model.
Clin Microbiol Infect, 15(7):651-655.
[3] Brandsberg, J.W., French, M.E., 1972.
In vitro susceptibility of isolates of
Aspergillus fumigatus and
Sporothrix schenckii to amphotericin B.
Antimicrob Agents Chemother, 2(5):402-404.
[4] Cruz, R., Vieille, P., Oschilewski, D., 2012.
Sporothrix globosa isolation related to a case of lymphocutaneous sporotrichosis.
Rev Chil Infectol, 29(4):401-405.
[5] de Beer, Z.W., Harrington, T.C., Vismer, H.F., 2003. Phylogeny of the
Ophiostoma stenoceras-
Sporothrix schenckii complex.
Mycologia, 95(3):434-441.
[6] de Lima Barros, M.B., de Oliveira Schubach, A., do Valle, A.C.F., 2004. Cat-transmitted sporotrichosis epidemic in Rio de Janeiro, Brazil: description of a series of cases.
Clin Infect Dis, 38(4):529-535.
[7] de Oliveira, M.M.E., de Almeida-Paes, R., de Medeiros Muniz, M., 2010. Sporotrichosis caused by
Sporothrix globosa in Rio de Janeiro, Brazil: case report.
Mycopathologia, 169(5):359-363.
[8] Fernandes, G.F., dos Santos, P.O., Rodrigues, A.M., 2013. Characterization of virulence profile, protein secretion and immunogenicity of different
Sporothrix schenckii sensu stricto isolates compared with
S. globosa and
S. brasiliensis species.
Virulence, 4(3):241-249.
[9] Galhardo, M.C.G., Silva, M.T.T., Lima, M.A., 2010.
Sporothrix schenckii meningitis in AIDS during immune reconstitution syndrome.
J Neurol Neurosurg Psychiatry, 81(6):696-699.
[10] Gilgado, F., Serena, C., Cano, J., 2006. Antifungal susceptibilities of the species of the
Pseudallescheria boydii complex.
Antimicrob Agents Chemother, 50(12):4211-4213.
[11] Glass, N.L., Donaldson, G.C., 1995. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes.
Appl Environ Microbiol, 61(4):1323-1330.
[12] Hall, T.A., 1999. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT.
Nucl Acids Symp Series, 41:95-98.
[13] Katoh, K., Toh, H., 2010. Parallelization of the MAFFT multiple sequence alignment program.
Bioinformatics, 26(15):1899-1900.
[14] Kong, X., Xiao, T., Lin, J., 2006. Relationships among genotypes, virulence and clinical forms of
Sporothrix schenckii infection.
Clin Microbiol Infect, 12(11):1077-1081.
[15] Koufopanou, V., Burt, A., Szaro, T., 2001. Gene genealogies, cryptic species, and molecular evolution in the human pathogen
Coccidioides immitis and relatives (
Ascomycota,
Onygenales).
Mol Biol Evol, 18(7):1246-1258.
[16] Lin, J., Kawasaki, M., Aoki, M., 2000. Mitochondrial DNA analysis of
Sporothrix schenckii clinical isolates from China.
Mycopathologia, 148(2):69-72.
[17] Lopes-Bezerra, L.M., Schubach, A., Costa, R.O., 2006.
Sporothrix schenckii and sporotrichosis.
An Acad Bras Ciênc, 78(2):293-308.
[18] Lpez-Romero, E., del Roco Reyes-Montes, M., Prez-Torres, A., 2011.
Sporothrix schenckii complex and sporotrichosis, an emerging health problem.
Future Microbiol, 6(1):85-102.
[19] Madrid, H., Cano, J., Gen, J., 2009.
Sporothrix globosa, a pathogenic fungus with widespread geographical distribution.
Rev Iberoam Micol, 26(3):218-222.
[20] Madrid, H., Gen, J., Cano, J., 2010.
Sporothrix brunneoviolacea and
Sporothrix dimorphospora, two new members of the
Ophiostoma stenoceras-
Sporothrix schenckii complex.
Mycologia, 102(5):1193-1203.
[21] Marimon, R., Gen, J., Cano, L., 2006. Molecular phylogeny of
Sporothrix schenckii
.
J Clin Microbiol, 44(9):3251-3256.
[22] Marimon, R., Cano, J., Gen, J., 2007.
Sporothrix brasiliensis,
S. globosa, and
S. mexicana, three new
Sporothrix species of clinical interest.
J Clin Microbiol, 45(10):3198-3206.
[23] Marimon, R., Serena, C., Gen, J., 2008.
In vitro antifungal susceptibilities of five species of
Sporothrix
.
Antimicrob Agents Chemother, 52(2):732-734.
[24] Marimon, R., Gen, J., Cano, J., 2008.
Sporothrix luriei: a rare fungus from clinical origin.
Med Mycol, 46(6):621-625.
[25] McGinnis, M., Nordoff, N., Li, R.K., 2001.
Sporothrix schenckii sensitivity to voriconazole, itraconazole and amphotericin B.
Med Mycol, 39(4):369-371.
[26] Mei, X.L., Xia, J.X., Wang, J.Y., 2011. Clinical and pathological analysis on 100 cases of sporotrichosis.
Chin J Mycol, (in Chinese),6:203-206.
[27] Mesa-Arango, A.C., del Roco Reyes-Montes, M., Prez-Meja, A., 2002. Phenotyping and genotyping of
Sporothrix schenckii isolates according to geographic origin and clinical form of sporotrichosis.
J Clin Microbiol, 40(8):3004-3011.
[28] Nylander, J., 2004. MrModeltest V2. Program Distributed by the Author. Evolutionary Biology Centre,Uppsala University :
[29] Page, R.D., 1996. TreeView: an application to display phylogenetic trees on personal computers.
Comput Appl Biosci, 12(4):357-358.
[30] Porebski, S., Bailey, L.G., Baum, B.R., 1997. Modification of a CTAB DNA extraction protocol for plants containing high polysaccharide and polyphenol components.
Plant Mol Biol Rep, 15(1):8-15.
[31] Rodrigues, A.M., de Hoog, S., de Camargo, Z.P., 2012. Emergence of pathogenicity in the
Sporothrix schenckii complex.
Med Mycol, 51(4):405-412.
[32] Schoch, C.L., Seifert, K.A., Huhndorf, S., 2012. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for fungi.
PNAS, 109(16):6241-6246.
[33] Schubach, A., de Lima Barros, M.B., Wanke, B., 2008. Epidemic sporotrichosis.
Curr Opin Infect Dis, 21(2):129
[34] Silva-Vergara, M.L., de Camargo, Z.P., Silva, P.F., 2012. Disseminated
Sporothrix brasiliensis infection with endocardial and ocular involvement in an HIV-infected patient.
Am J Tropical Med Hygiene, 86(3):477
[35] Song, Y., Yao, L., Zhong, S.X., 2011. Infant sporotrichosis in northeast China: a report of 15 cases.
Int J Dermatol, 50(5):522-529.
[36] Song, Y., Li, S.S., Zhong, S.X., 2011. Report of 457 sporotrichosis cases from Jilin Province, northeast China, a serious endemic region.
J Eur Acad Dermatol Venereol, 27(3):313-318.
[37] Swofford, D.L., 2003. PAUP*. Phylogenetic Analysis Using Parsimony (*and Other Methods). Version 4. Sinauer Associates,Sunderland, Massachusetts, USA :
[38] Tan, J.W., Liu, W., Wan, Z., 2013. Reclassification of 33 clinical strains of
Sporothrix from northern China based on phenotypic and molecular characters.
Mycosystema, (in Chinese),32(2):161-167.
[39] Watanabe, S., Kawasaki, M., Mochizuki, T., 2004. RFLP analysis of the internal transcribed spacer regions of
Sporothrix schenckii
.
Jpn J Med Mycol, 45:165-175.
[40] White, T.J., Bruns, T., Lee, S., 1990. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics.
PCR Prot A Guide Meth Appl, 18:315-322.
[41] Yap, F.B.B., 2011. Disseminated cutaneous sporotrichosis in an immunocompetent individual.
Int J Infect Dis, 15(10):727-729.
[42] Zhang, J.D., Lin, J.P., 2008. Clinical analysis of 316 cases of cutaneous sporotrichosis.
Chin J Mycol, (in Chinese),3:207-210.
[43] Zhou, X., Rodrigues, A.M., Feng, P., 2013. Global ITS diversity in the
Sporothrix schenckii complex.
Fungal Div, in press,:

Open peer comments: Debate/Discuss/Question/Opinion
<1>