1. Introduction
5-Aminolevulinic acid (ALA) is an endogenously synthesized amino acid involved in the synthesis of tetrapyrroles such as vitamin B
12, heme, and chlorophyll
in vivo. It is a precursor of tetrapyrroles and a key intermediate in regulating tetrapyrrole biosynthesis (Sasaki et al.,
2002). ALA has been used in a number of applications, i.e., as a new kind of green pesticide in agriculture and as a photodynamic medicine for photodynamic diagnosis and therapy of cancer (Lang et al.,
2001; Braathen et al.,
2007; Berkovitch et al.,
2008; Pariser et al.,
2008; Zaak et al.,
2008).
Compared with conventional methods, microbial production of ALA with new technologies such as genetic engineering and metabolic engineering has recently attracted much attention. Recombinant bacteria containing the
hemA gene from such organisms as
Rhodobacter sphaeroides (Vanderwerf and Zeikus,
1996; Kiatpapan and Murooka,
2001; Xie et al.,
2003b; Wang et al.,
2006; Fu et al.,
2007),
Bradyrhizobium japonicum (Choi et al.,
1999; Lee et al.,
2004),
Rhodopseudomonas palustris (Choi et al.,
2008), and
Agrobacterium radiobacter (Liu et al.,
2005; Lin et al.,
2009) were constructed for ALA production, achieving levels of 39 mmol/L (Xie et al.,
2003a), 50 mmol/L (Fu et al.,
2008), and 56 mmol/L (Lin et al.,
2009), respectively.
In our previous work, ALA production by recombinant
Escherichia coli containing the
hemA gene from
R. sphaeroides (Fu et al.,
2007;
2008) and
A. radiobacter zju-0121 (Lin et al.,
2009; Fu et al.,
2010) reached a relatively high level. However, it has been difficult to achieve a substantial breakthrough since our work mainly focused on the optimization of fermentation conditions. In this study, we investigated the difference in the
hemA genes from different organisms as a starting point to obtain 5-aminolevulinic acid synthase (ALAS) with excellent enzymatic properties, which has high-level solubility expressed in recombinant
E. coli, resulting in further enhancement of ALA production.
Rhodobacter capsulatus is a good source of the
hemA gene. The gene was successfully cloned and expressed under the control of a constitutive promoter in
E. coli, and the recombinant
E. coli strain was able to accumulate ALA at 21 mmol/L (Kang et al.,
2004). However, the enzymatic properties remain unknown. In this study, the
R. capsulatus hemA gene was highly expressed in
E. coli Rosetta (DE3), which is a rare codon optimizer host strain. The recombinant ALAS (RC-ALAS) was isolated and purified, and its enzymatic properties were investigated.
Since the methods and analytical instruments for the determination of enzyme activity vary in different reports, the enzymatic properties are likely to be different, even if the enzyme was encoded by the same
hemA gene. For example, the activity of RS-ALAS, which was encoded by the
hemA gene from
R. sphaeroides, was reported to be 130 μmol/(h∙mg) (Warnick and Burnham,
1971), while it was found to be 13 μmol/(h∙mg) by Bolt et al. (
1999). The activity of RP-ALAS, encoded by the
hemA gene from
Rhodopseudomonas palustris, has been reported as 65.23 nmol/(h∙mg) (Viale et al.,
1987) and 27.73 μmol/(30 min∙mg) (Choi et al.,
2004), a difference of three orders of magnitude. To avoid this problem, the enzymatic properties of purified RC-ALAS were investigated under the same conditions for AR-ALAS and RS-ALAS, which were encoded by the
hemA gene from
A. radiobacter zju-0121 and
R. sphaeroides, respectively.
2. Materials and methods
2.1. Bacterial strains, plasmids, and growth conditions
The bacterial strains and plasmids used in this study are listed in Table
1.
Table 1
Bacterial strains and plasmids used in this study
| Strain or plasmid |
Relevant property |
Source |
| Strain |
| Rhodobacter capsulatus ATCC 11166 |
Type strain |
DSMZ |
| E. coli Rosetta (DE3)/pET28a-A.R.hemA
|
CmR & KanR
|
Laboratory stock |
| E. coli Rosetta (DE3)/pET28a-R.S.hemA
|
CmR & KanR
|
Laboratory stock |
| E. coli Rosetta (DE3) |
CmR
|
Laboratory stock |
| Plasmid |
| pET28a(+) |
KanR
|
Novagen |
CmR: chloramphenicol resistance; KanR: kanamycin resistance; DSMZ: Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH
strains were cultivated in Luria-Bertani (LB) medium supplemented, when necessary, with kanamycin (30 mg/L) or chloramphenicol (34 mg/L).
R. capsulatus was cultivated under anaerobic conditions in the light at 25 °C, using modified Rhodospirillaceae medium containing 0.30 g/L yeast extract, 1.00 g/L sodium succinate, 0.50 g/L ammonium acetate, 5.00 ml Fe(III) citrate solution (1 g/L in H
2O), 0.50 g/L KH
2PO
4, 0.40 g/L MgSO
4∙7H
2O, 0.40 g/L NaCl, 0.40 g/L NH
4Cl, 0.05 g/L CaCl
2∙2H
2O, 0.40 ml vitamin B
12 solution (10 mg in 100 ml H
2O), 1.00 ml trace element solution SL-6 (0.10 g/L ZnSO
4∙7H
2O, 0.03 g/L MnCl
2∙4H
2O, 0.30 g/L H
3BO
3, 0.20 g/L CoCl
2∙6H
2O, 0.01 g/L CuCl
2∙2H
2O, 0.02 g/L NiCl
2∙6H
2O, and 0.03 g/L Na
2MoO
4∙2H
2O), 0.3 g/L L-cysteine chloride, and 0.50 ml resazurin (1 g/L in H
2O).
2.2. Construction of the expression plasmid containing the hemA gene from R. capsulatus
The genomic DNA of
R. capsulatus was extracted by using a Bacterial Genomic DNA Miniprep Kit (Axygen). Two oligonucleotide primers, ALA1 (5′-GGGAATTC
CATATGGACTACAATCTCGCGCTC-3′) and ALA2 (5′-CCC
AAGCTTAGGCCTCGGCGCGATTCAC-3′), which were designed with
NdeI and
HindIII restriction sites (underlined), respectively, were synthesized for the cloning of the
hemA gene from the genomic DNA by polymerase chain reaction (PCR) amplification. The amplification was conducted in a 50-μl reaction mixture containing primers (10 pmol), genomic DNA (50 ng), 10 μl of 5× PrimeSTAR GXL buffer, 4 μl of dNTP mixture (2.5 mmol/L each), and 1 μl of PrimeSTAR GXL DNA polymerase (TaKaRa). The reaction mixture was amplified for 30 cycles of denaturation at 98 °C for 10 s, annealing at 58 °C for 15 s, and extension at 72 °C for 80 s.
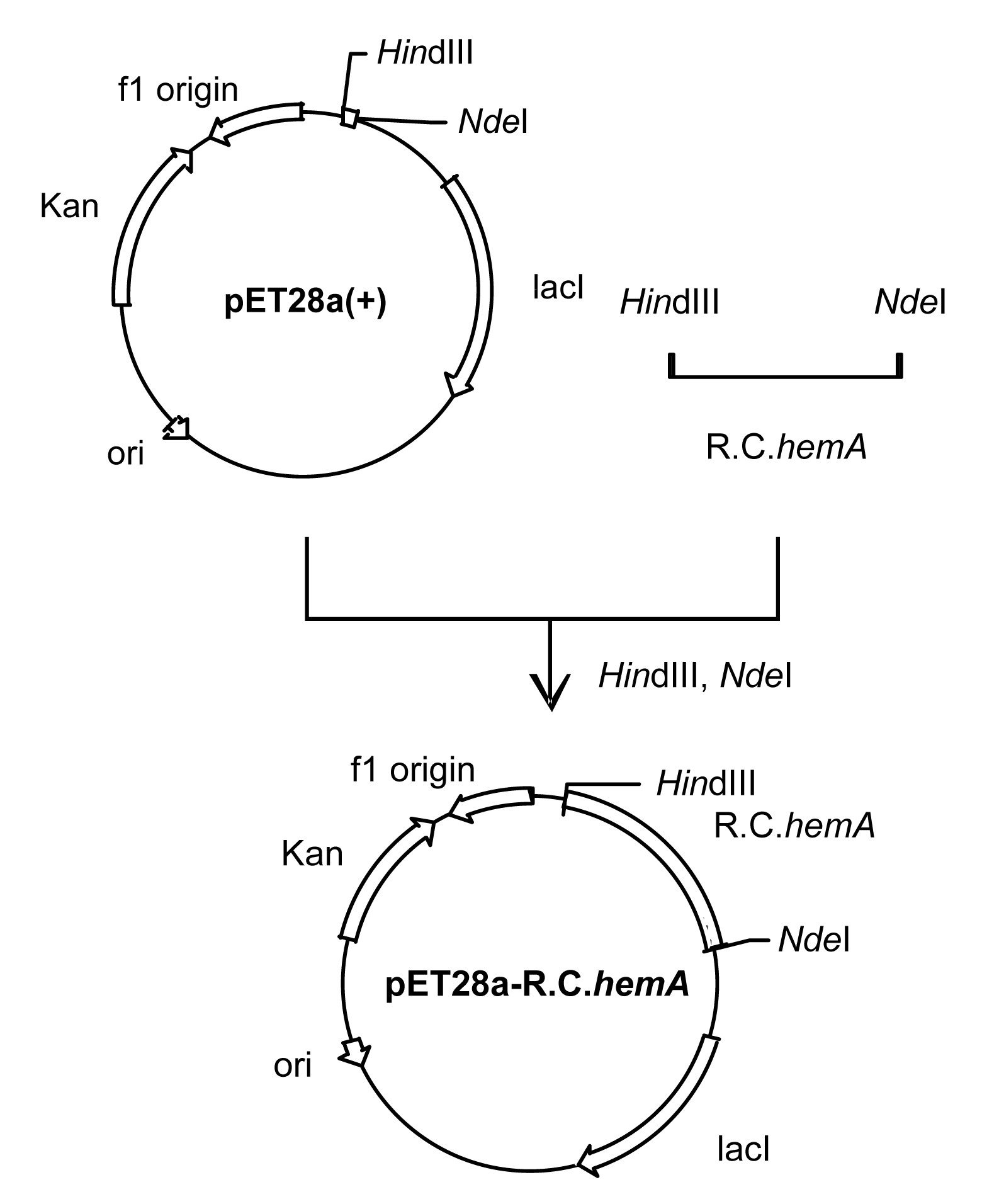
The amplified 1.2-kb DNA fragment was isolated and cloned into the pET28a(+) expression vector (Fig.
1). The resulting plasmid, designated pET28a-R.C.
hemA, was transformed into
E. coli Rosetta (DE3) to obtain
E. coli Rosetta (DE3)/pET28a-R.C.
hemA.
Fig.1
Construction of expression vector
2.3. Expression of recombinant hemA gene and purification of ALAS
The three recombinant ALASs, RC-ALAS, AR-ALAS, and RS-ALAS, were expressed and purified using the method described in our previous work (Lin et al.,
2009). Specifically, for the cultivation of recombinant
E. coli, LB medium supplemented with kanamycin (30 mg/L) and chloramphenicol (34 mg/L) was used. After incubation for 2 h at 37 °C, isopropyl-β-D-thiogalactopyranoside (IPTG) was added to induce the expression of ALAS at the final concentration of 0.05 mmol/L, and the mixture was incubated for 6 h at 28 °C. Cells were harvested by centrifugation (10 000×
g for 10 min at 4 °C). The cells were resuspended in 50 mmol/L Tris-buffer (pH 7.5), and then disrupted with an ultrasonicator (Ningbo Scientz Biotechnology, China) for 6 min. After re-centrifugation (10 000×
g for 10 min at 4 °C), the 6-His-tagged ALAS in the supernatant was recovered by affinity purification on nickel-nitrilotriacetic acid (Ni-NTA) according to the manufacturer’s instructions (Qiagen, Germany), and was eluted with imidazole and NaCl. Sephadex G-25 medium resin (GE Healthcare, USA) was used for gel filtration chromatography to remove imidazole and NaCl from the ALAS solution to obtain purified ALAS. The purity of ALAS was determined by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) analysis.
2.4. Assay for enzyme activity and ALA
Protein concentrations were measured using a Pierce BCA protein assay kit (Sigma, St. Louis, MO, USA) with bovine serum albumin as the standard. The enzyme assay was carried out according to our previous work (Lin et al.,
2009) with slight modification. The assays were conducted in a 500-μl reaction mixture containing 380 μl of about 0.1 mg/ml purified ALAS and 120 μl of reaction liquid containing 50 μl of 1 mol/L glycine, 50 μl of 2 mmol/L succinyl-CoA, 6 μl of 1 mol/L Tris-buffer (pH 7.5), and 14 μl of 10 mmol/L pyridoxal phosphate, which were mixed together after preheating at 37 °C for 5 min, respectively. After proceeding at 37 °C for 10 min, the reaction was terminated by the addition of 150 μl of 10% (v/v) trichloroacetic acid. Then, the sample was centrifuged and assayed to determinate the concentration of ALA in the supernatant. One unit of ALAS activity was defined as the amount of enzyme needed to produce 1 nmol of ALA in 1 min. The ALA concentration was determined according to our previous work (Fu et al.,
2008). Specifically, 2 ml of sample or standard was mixed with 1 ml 1.0 mol/L sodium acetate (pH 4.6) in a cuvette. Then the mixtures were heated at 100 °C for 15 min. After cooling for 15 min, 2 ml of reaction mixture and 2 ml freshly prepared modified Ehrlich’s reagent (Burnham,
1970) were mixed together. After 30 min, the absorbance at 554 nm was measured.
2.5. Enzyme properties of ALAS
The enzymatic properties of purified RC-ALAS, AR-ALAS, and RS-ALAS were studied under the same conditions. The enzymatic reactions were assayed at various pH values and temperatures to determine the optimum pH and temperature, as well as pH and temperature stability. The effects of various metal ions, SDS, and ethylene diamine tetraacetic acid (EDTA) on enzyme activity were also evaluated. All the measurements were performed with 0.1 mg/ml ALAS in 50 mmol/L Tris-HCl (pH 7.5) buffer. The glycine concentration was set at 100 mmol/L and different concentrations of succinyl-CoA were used to examine the relationship between the initial velocity and the substrate concentration. The kinetic parameters, Michaelis constant (
K
m) and maximal velocity (
V
max), were determined by fitting the data to the Michaelis-Menten equation using non-linear regression analysis software (SigmaPlot10; Systat).
2.6. Fermentation conditions
Seed cultures were incubated for 3 h at 200 r/min and at 37 °C in shaken flasks containing 200 ml LB medium supplemented with kanamycin (30 mg/L) and chloramphenicol (34 mg/L). The fermentation was conducted in a 15-L fermenter (Shanghai Guoqiang Bioengineering Equipment Co., Ltd., China). Nine-liter sterilized fermentation medium containing 20 g/L glycerol, 10 g/L yeast extract, 2 g/L glycine, and 3 g/L succinic acid was added, and 400 ml seed culture was inoculated to provide an initial optical density at 600 nm (OD
600) of approximately 0.1. The fermenter was operated at 400 r/min and the air flow rate was regulated at 6 L/min. After cultivation at 37 °C for 2 h, IPTG was added and the temperature was lowered to 28 °C. During the fermentation, the feed addition was performed by a peristaltic pump. The feed stream was a mixture of precursors containing 75 g/L succinic acid and 65 g/L glycine, and the feeding rate was regulated by the pH feed-back control system to maintain the pH value.
3. Results and discussion
3.1. Cloning and expression of the R. capsulatus hemA gene
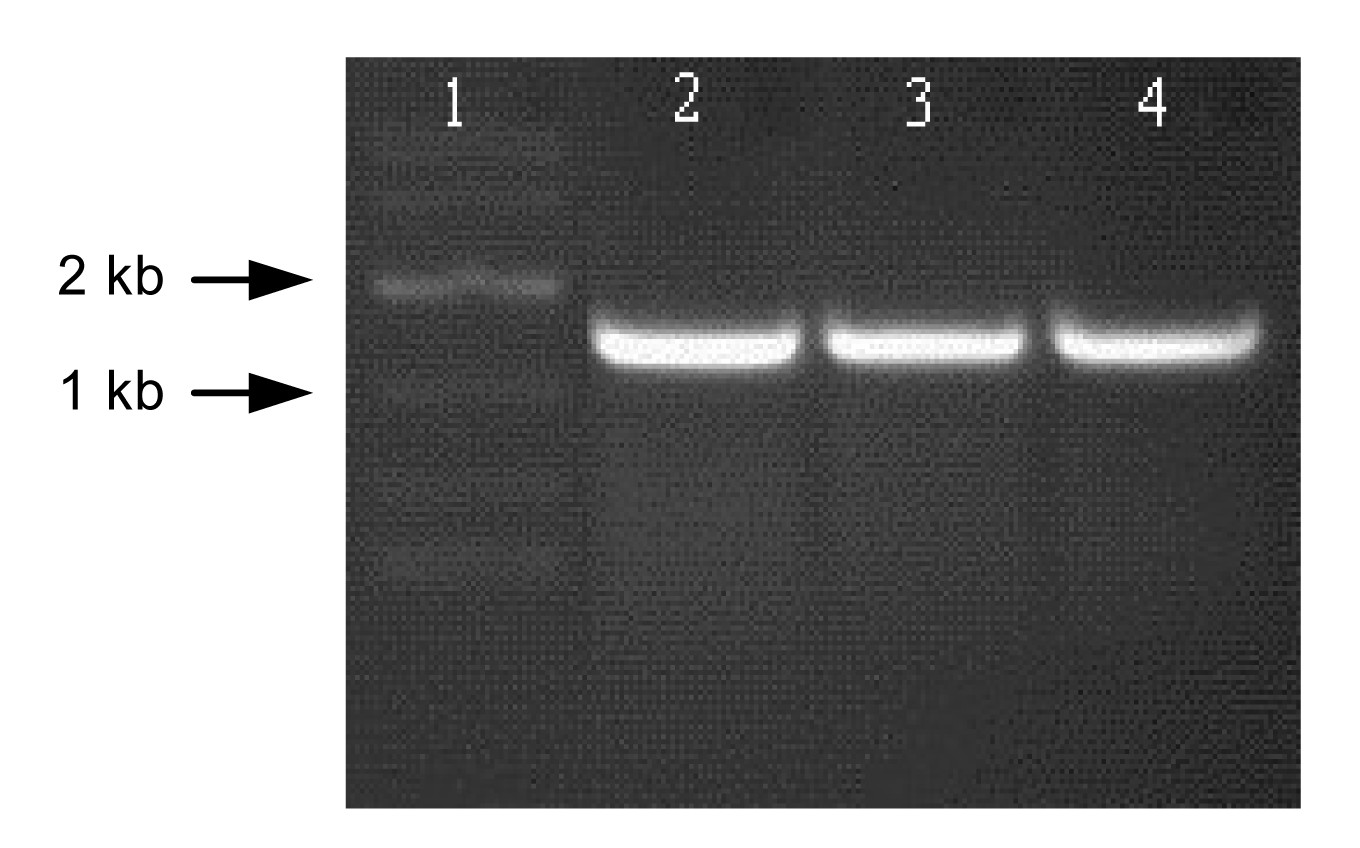
As shown in Fig.
2, a 1.2-kb DNA fragment carrying
hemA gene was isolated by PCR from genomic DNA of
R. capsulatus. Sequencing experiments showed that the
hemA coding region was 1206 bp long, indicating a protein of 402 amino acids with a molecular weight of about 44 kD. The DNA sequence showed 77% homology with the
A. radiobacter zju-0121
hemA gene (GeneID: 88174324) and 64% homology with the
R. sphaeroides hemA gene (GeneID: 3720398). Meanwhile, the amino acid sequence of RC-ALAS shared 58% identity with AR-ALAS and 75% identity with RS-ALAS, which suggested that the enzymatic properties of the three ALASs might be different.
Fig.2
Agarose gel electrophoresis analysis of hemA from R. capsulatus
Lane 1: marker; Lanes 2–4: hemA
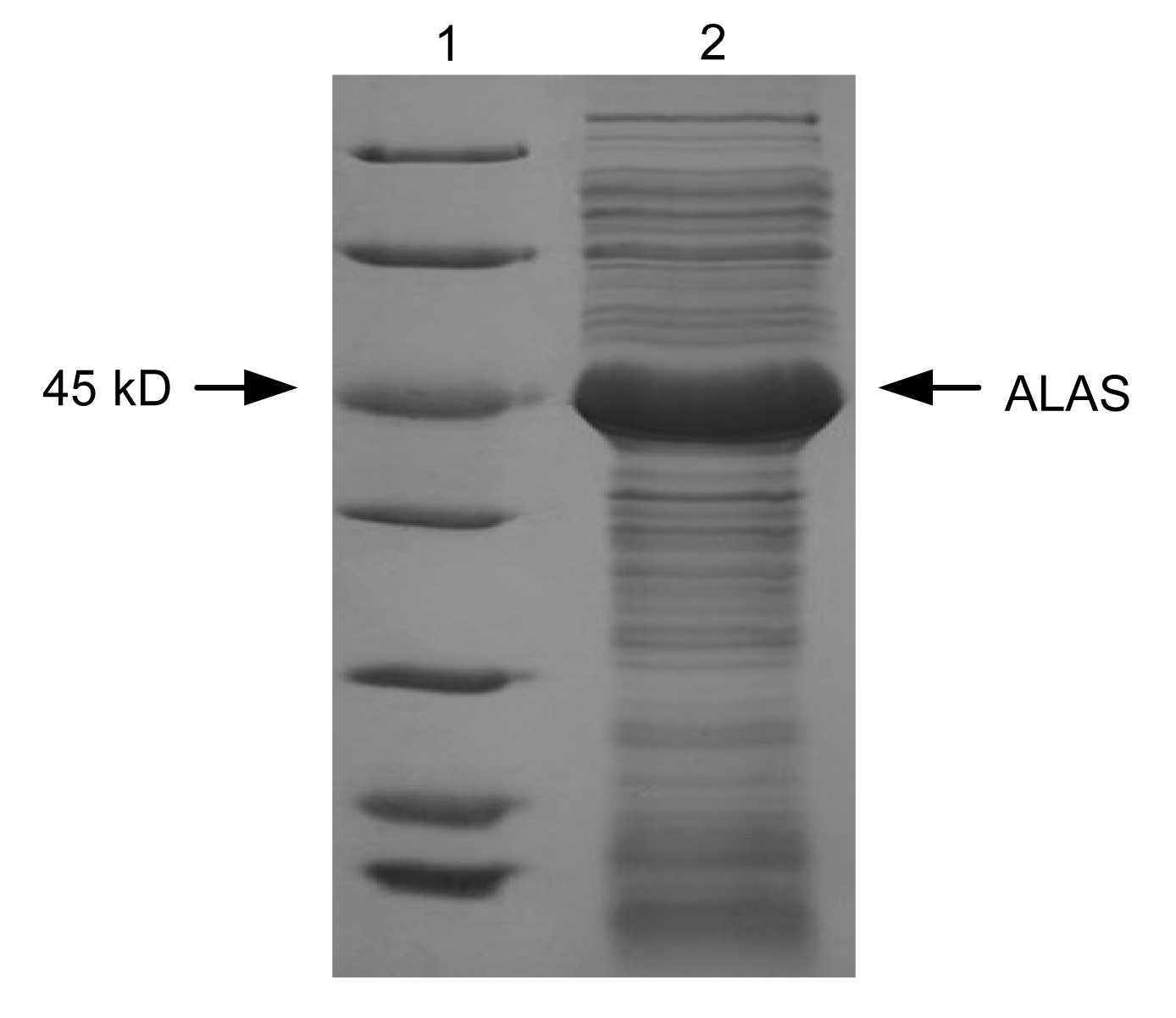
The SDS-PAGE analysis of the recombinant RC-ALAS, which was highly expressed by
E. coli Rosetta (DE3)/pET28a-R.C.
hemA, is shown in Fig.
3.
Fig.3
Expression of the R. capsulatus hemA gene inE. coli Rosetta (DE3)
Lane 1: marker; Lane 2: crude RC-ALAS
3.2. Enzymatic properties of recombinant ALAS
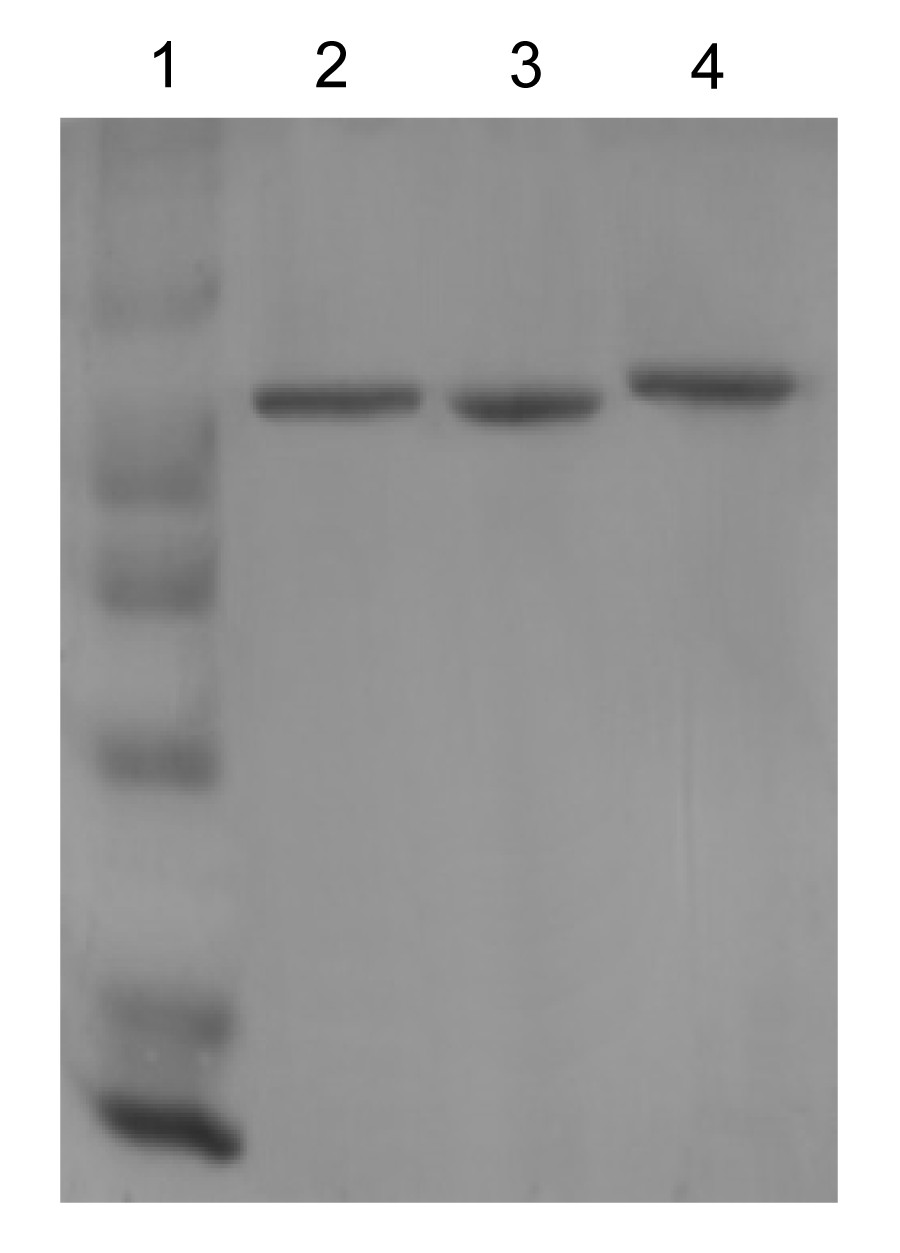
The purified recombinant proteins, RC-ALAS, AR-ALAS, and RS-ALAS, were obtained after Ni-NTA agarose affinity purification and Sephadex G-25 medium resin gel filtration chromatography, and the purity was determined by SDS-PAGE with about 0.1 mg/ml ALAS, as shown in Fig.
4. The enzymatic properties of all three were studied under the same conditions.
Fig.4
SDS-PAGE analysis of purified recombinant ALASs
Lane 1: marker; Lane 2: RC-ALAS; Lane 3: AR-ALAS; Lane 4: RS-ALAS
3.2.1. Optimum temperature
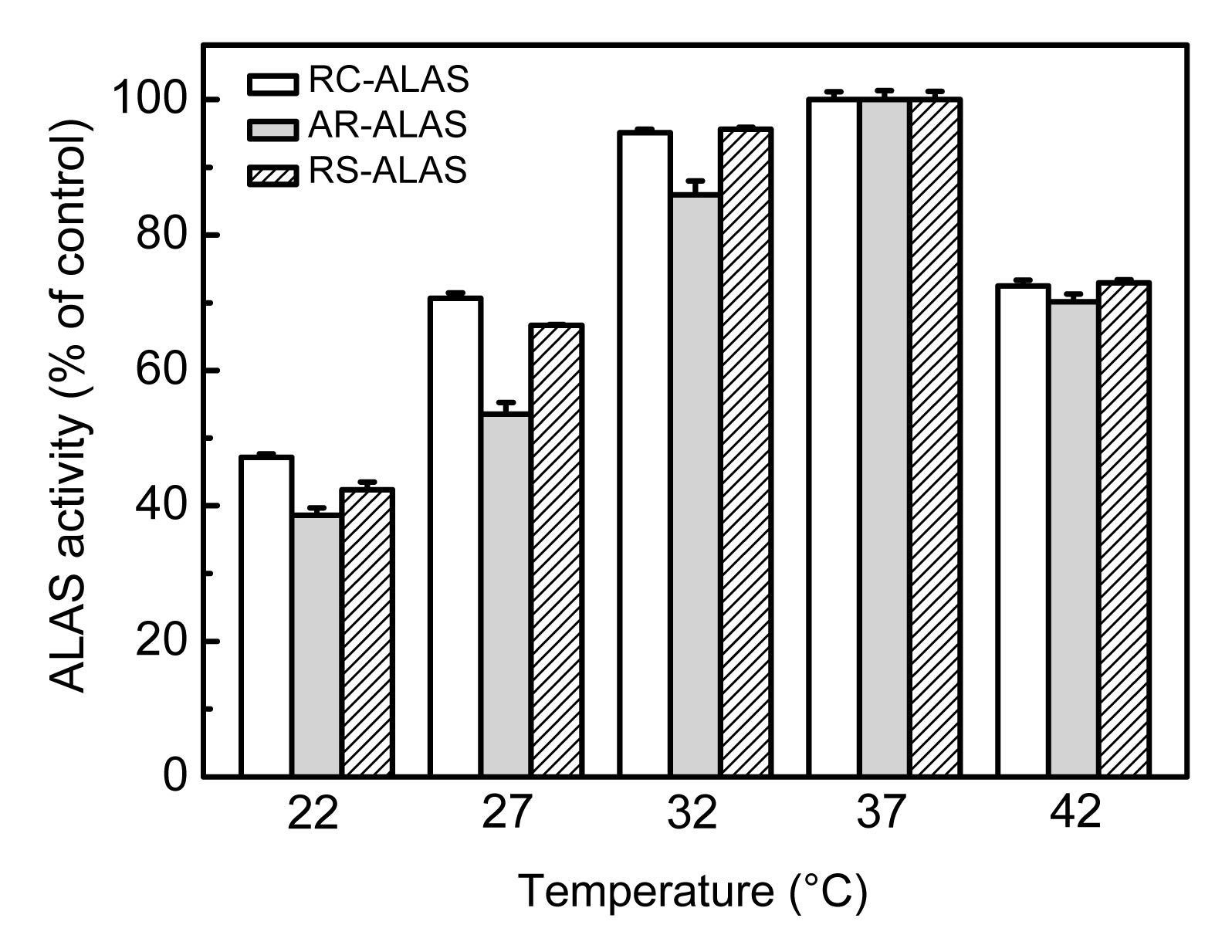
The enzymatic reactions were assayed at various temperatures ranging from 22 to 42 °C. As shown in Fig.
5, the optimum temperature of all three enzymes was found to be 37 °C. Among the three, a marked effect of temperature was observed for AR-ALAS, while a minimal impact was found for RC-ALAS. In particular, when the temperature was lowered from 37 to 22 °C, the specific activities of the three enzymes dropped to below 50%. Moreover, when the temperature was increased to 42 °C, a significant decrease in enzyme activity was observed. It was concluded that the fermentation temperature could directly influence the enzyme activity. This result suggests that we need to consider the effects of temperature on the growth of recombinant bacteria, as well as the expression and specific activity of the recombinant enzyme as a whole during fermentation.
Fig.5
Effects of various temperatures on the activity of recombinant ALAS
The activity of the sample assayed at 37 °C was set as 100%. Data are expressed as mean±SD (n=9)
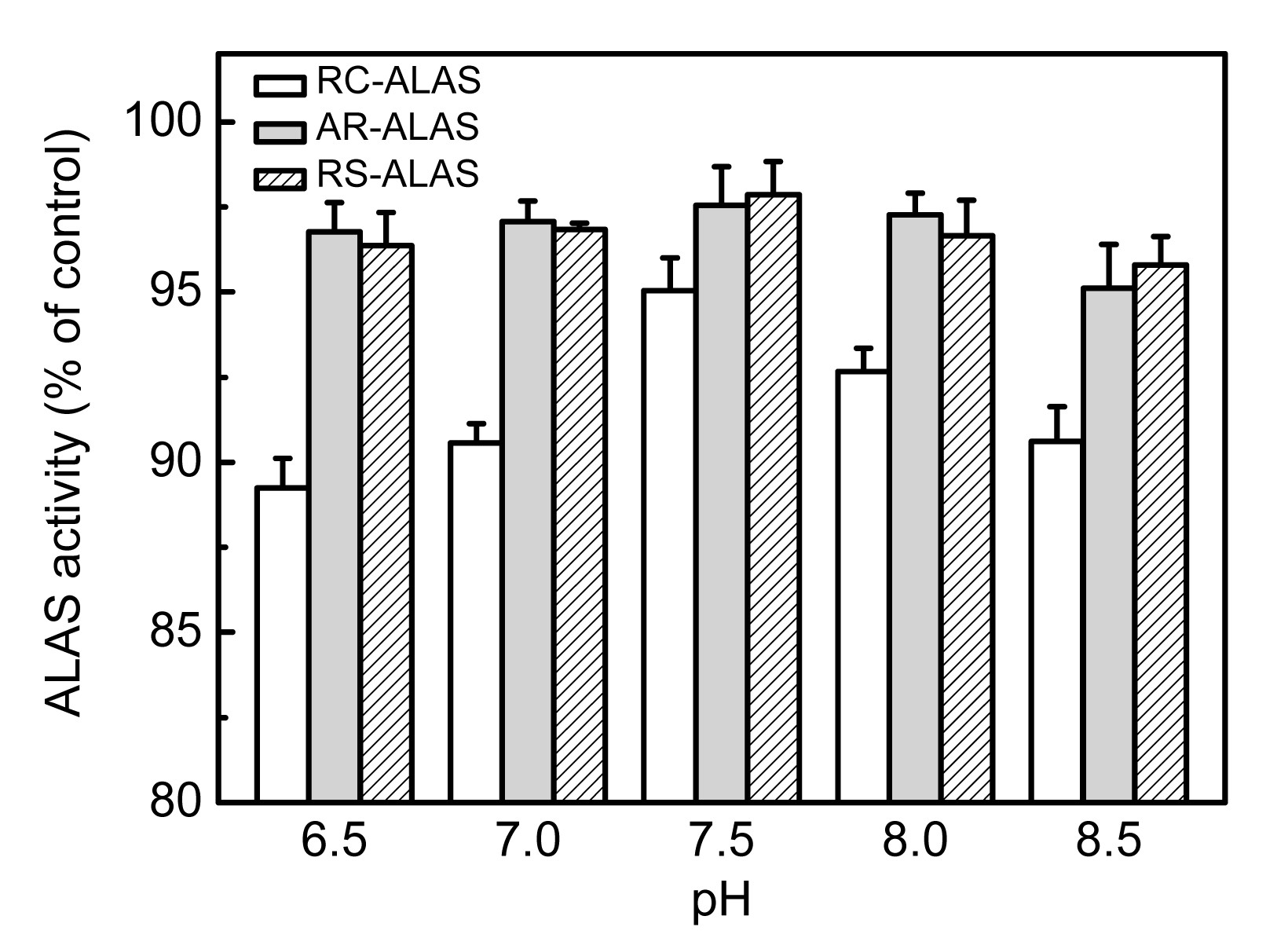
3.2.2. Optimum pH
The enzymatic reactions were assayed at various pH values ranging from 6.5 to 8.5. As shown in Fig.
6, the optimum pH value of all three enzymes was 7.5. Acid effectively inhibited the activity of RC-ALAS, since its activity dropped to below 80%, while the other two remained above 90% at pH 6.5. Conversely, strong alkaline conditions exerted the greatest inhibition of the activity of AR-ALAS, and had a minimal impact on RS-ALAS.
Fig.6
Effects of various pH values on the activity of recombinant ALAS
The activity of the sample assayed at pH 7.5 was set as 100%. Data are expressed as mean±SD (n=9)
According to previous work (Fan,
2006), ALA was found to be unstable in aqueous solution, which was dependent on various parameters such as pH and temperature. ALA degraded with increased pH, and it was relatively stable in acidic conditions. Generally, ALA production was carried out in slightly acidic conditions, owing to its instability in alkaline conditions and difficulties in applications with strongly acidic conditions. As shown in Fig.
6, a nearly 20% decline in the activity of RC-ALAS was observed when the pH decreased from 7.0 to 6.5, indicating that a slight increase of the initial medium pH value may result in greater ALA production. This result suggests that the control of pH during fermentation is particularly important for the recombinant strain
E. coli Rosetta (DE3)/pET28a-R.C.
hemA.
3.2.3. Enzyme activity
The specific activity was assayed at the optimum temperature of 37 °C and optimum pH of 7.5. As listed in Table
2, the activity of RC-ALAS reached 198.2 U/mg, which was about 31.2% and 69.5% higher than those of AR-ALAS and RS-ALAS, respectively. Currently, the recombinant strain
E. coli Rosetta (DE3)/pET28a-A.R.
hemA is used in industrial mass production. Furthermore, the recombinant strain
E. coli Rosetta (DE3)/pET28a-R.C.
hemA may achieve much greater ALA production via the optimization of fermentation conditions, indicating its potential prospects for industrial application.
Table 2
Activity of recombinant ALAS
| ALAS |
Activity (U/mg)
|
| Crude enzyme |
Purified enzyme |
| RC-ALAS |
47.0 |
198.2 |
| AR-ALAS |
19.5 |
151.1 |
| RS-ALAS |
33.6 |
116.9 |
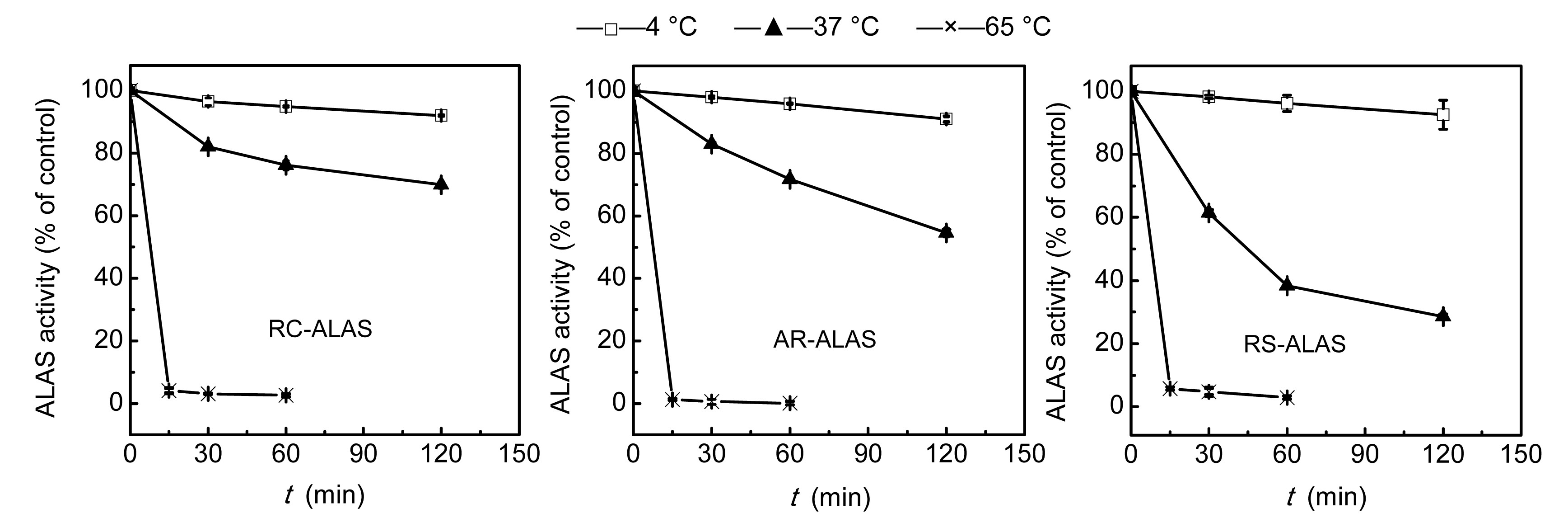
3.2.4. Temperature and pH stability
The temperature and pH stabilities of the three enzymes are shown in Figs.
7 and
8, respectively. After storage for 60 min at 65 °C, the three enzymes lost all their activity. After storage for 120 min at 37 °C, RS-ALAS retained only about 30% of its activity, while AR-ALAS retained about 50%. However, about 90% of activity remained after storage for 120 min at 4 °C. It is clear, therefore, that ALAS should be stored at a low temperature. All three enzymes were stable in the pH range 6.5 to 8.5 with all activities retained about or above 90%. In particular, RC-ALAS was more sensitive to pH, while the other two were sensitive to temperature.
Fig.7
Temperature stability of recombinant ALAS
Data are expressed as mean±SD (n=3)
Fig.8
pH stability of recombinant ALAS
Data are expressed as mean±SD (n=3)
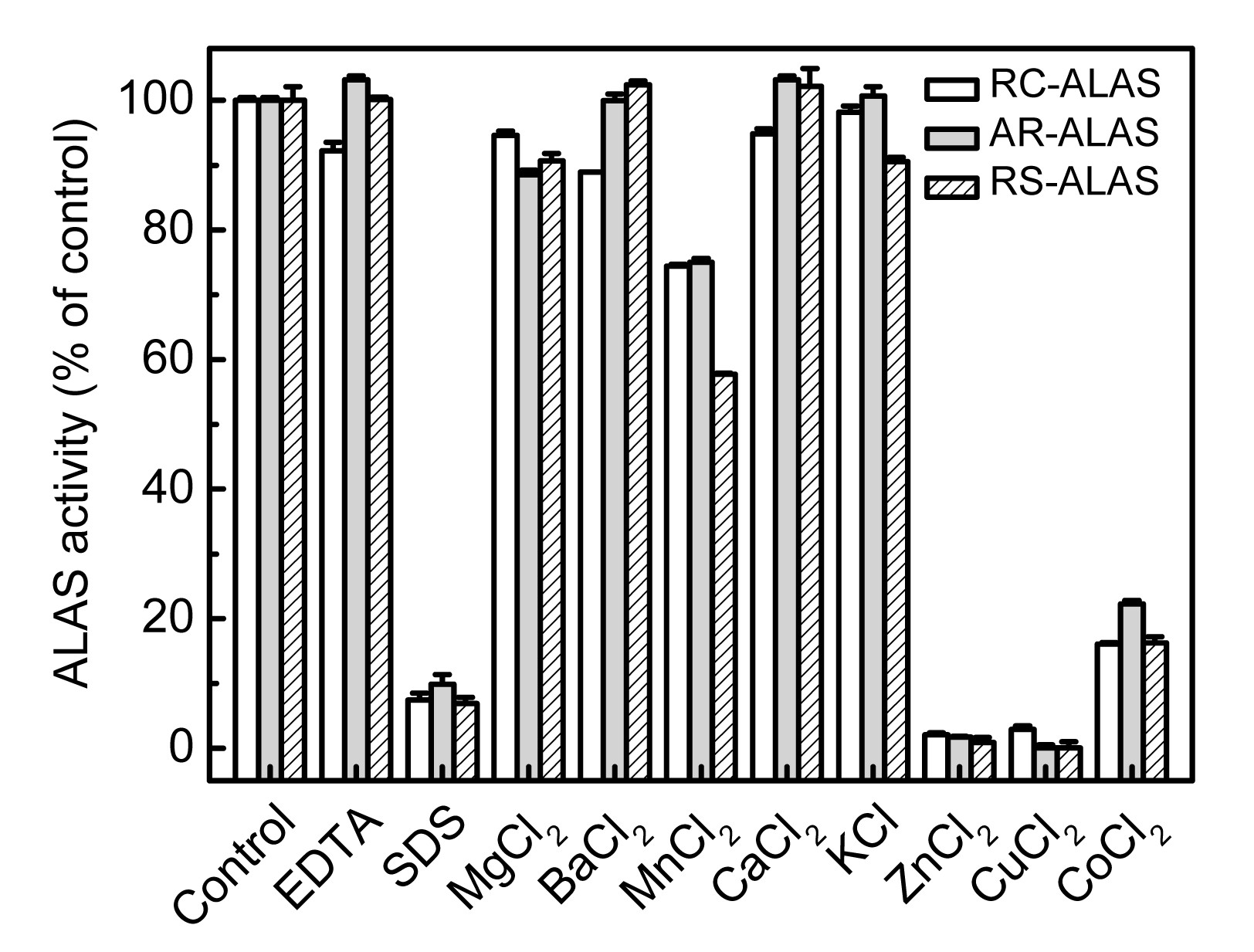
3.2.5. Effects of metal ions, EDTA, and SDS
In many cases, metal ions are bound to enzymes and play central roles in the catalytic process. Metal ions can function catalytically by forming nucleophiles, by direct coordination, by serving as electrophiles to stabilize a negative charge on a reactive intermediate, and by serving as a bridge between the enzyme and the substrate, which increases the binding energy. However, some heavy metals may cause enzyme denaturation.
The effects of various metal ions on enzyme activity were examined when the concentration of metal ions was set at 2 mmol/L. As shown in Fig.
9, the effects on the activities of the three enzymes were almost the same. Metal ions such as Co
2+, Zn
2+, and Cu
2+ strongly inhibited the activity of ALAS, while K
+, Ca
2+, Ba
2+, or Mg
2+ had no significant effect. Mn
2+ slightly inhibited the activity of ALAS, contrary to the observation in our previous work (Lin et al.,
2009). This difference is probably attributable to the fact that, in our previous work, we used a higher concentration of metal ions, thus exceeding the buffering capacity of the 50 mmol/L Tris-HCl buffer used.
Fig.9
Effects of various metal ions, EDTA, and SDS on the activity of recombinant ALAS
The activity of the untreated sample was set as 100%. Data are expressed as mean±SD (n=3)
As a metal ion chelator, EDTA can eliminate the interference caused by most of the metal ions, without affecting the function of the protein. We investigated whether EDTA influenced the activities of the three recombinant enzymes. The results show that EDTA had no significant effect on the activity of ALAS, indicating that metal ions are not required in the catalysis process.
SDS is an anionic detergent that can impart a negative charge or cause conformational changes to the active site of an enzyme, thus affecting its stability and activity. The results show that SDS significantly inhibited the activity of ALAS by as much as 90%.
3.2.6. Michaelis constant
The Michaelis constant,
K
m, is an important kinetic parameter of an enzyme. It is an inverse measure of the substrate’s affinity for the enzyme, so a small
K
m value indicates a high affinity. The turnover number,
k
cat, is the maximum number of substrate molecules converted to product per enzyme molecule per second. The ratio of
k
cat/
K
m is often used as a measure of enzyme efficiency.
ALAS catalyzes the condensation of glycine and succinyl-CoA to produce ALA, in which the substrates are glycine and succinyl-CoA. However, in the actual production, succinyl-CoA is replaced by succinic acid, which is often used as a precursor that can transform reversibly to succinyl-CoA via the Krebs cycle; there are many inherent difficulties in the mass production and storage of succinyl-CoA. Meanwhile, glycine is often added in excess to enhance ALA production because of its low price, thus making succinyl-CoA the limiting factor of the reaction rate. Therefore, a higher affinity between ALAS and succinyl-CoA means better production of ALA.
The relationship between the initial velocity and the concentration of succinyl-CoA, ranging from 0.05 to 0.40 mmol/L, was examined while the concentration of glycine was set at 100 mmol/L. The kinetic parameters,
K
m and
V
max, were determined by fitting the data to the Michaelis-Menten equation using non-linear regression analysis software (SigmaPlot10). The turnover number
k
cat and the specificity constant of succinyl coenzyme A [(
k
cat/
K
m)
S-CoA] were calculated and shown in Table
3. The (
k
cat/
K
m)
S-CoA of RC-ALAS was 1.4989, which was higher than those of AR-ALAS and RS-ALAS, indicating its higher catalytic efficiency.
Table 3
Kinetic parameters of the enzymatic reaction of recombinant ALAS
| ALAS |
K
m
S-CoA (mmol/L) |
k
cat (s−1) |
(k
cat/K
m)S-CoA (L/(mmol∙s)) |
| RC-ALAS |
0.1986 |
0.2977 |
1.4989 |
| AR-ALAS |
0.2674 |
0.1994 |
0.7456 |
| RS-ALAS |
0.0764 |
0.0894 |
1.1699 |
3.3. ALA production by E. coli Rosetta (DE3)/pET28a-R.C.hemA
Based on the above results,
E. coli Rosetta (DE3)/pET28a-R.C.
hemA may be a high-yield strain for ALA production. It was therefore desirable to carry out the fermentation using
E. coli Rosetta (DE3)/pET28a-R.C.
hemA. In this work, a 15-L fermenter was used, and fed-batch fermentation carried out under the conditions that the above results suggest. The pH value was controlled at 6.2 initially and at 6.5 after 6 h of incubation with the pH feed-back controlled precursor feeding method. Meanwhile, the temperature was lowered to 28 °C after 2 h of incubation for the better expression of the
R. capsulatus hemA gene, and increased to 37 °C after 22 h to enhance the catalytic ability of RC-ALAS and the growth of the recombinant strain.
The results in Fig.
10 show that the yield of ALA achieved was 8.8 g/L (67 mmol/L) using the recombinant strain
E. coli Rosetta (DE3)/pET28a-R.C.
hemA under the appropriate conditions in the fermenter. This suggests that using the
hemA gene from
R. capsulatus increased the ALA yield over 20% and 33%, respectively, than the yields achieved by recombinant
E. coli containing the
hemA gene from
A. radiobacter zju-0121 (Lin et al.,
2009; Fu et al.,
2010) and
R. sphaeroides (Fu et al.,
2007;
2008).
Fig.10
Production of ALA using E. coli Rosetta (DE3)/pET28a-R.C.hemA
Mixture of glycine and succinate acid was added continuously by the pH feed-back control system. The pH value was controlled at 6.2 initially and at 6.5 after 6 h of incubation. The temperature was controlled at 28 °C after 2 h of incubation and increased to 37 °C after 22 h. Data are expressed as mean±SD (n=3)
4. Conclusions
The
hemA gene from
R. capsulatus (DSM1710) was highly expressed in
E. coli Rosetta (DE3), and its enzymatic properties were investigated in comparison with AR-ALAS and RS-ALAS. The activity of purified RC-ALAS reached 198.2 U/mg, which was about 31.2% and 69.5% higher than those of AR-ALAS (151.1 U/mg) and RS-ALAS (116.9 U/mg), respectively.
The results of the enzymatic properties suggest that we need to consider the effects of all the factors mentioned above on the growth of recombinant bacteria, as well as the expression and specific activity of the recombinant enzyme as a whole during fermentation to enhance the production of ALA. The fed-batch fermentation was conducted using recombinant strain
E. coli Rosetta (DE3)/pET28a-R.C.
hemA, and the yield of ALA achieved was 8.8 g/L (67 mmol/L), indicating a good potential for its industrial applications.
* Project supported by the National Natural Science Foundation of China (No. 20306026) and the National Basic Research Program (973) of China (No. 2007CB707805)Compliance with ethics guidelines Jia-wei LOU, Li ZHU, Mian-bin WU, Li-rong YANG, Jian-ping LIN, and Pei-lin CEN declare that they have no conflict of interest.References
[1] Berkovitch, G., Doron, D., Nudelman, A., 2008. Novel multifunctional acyloxyalkyl ester prodrugs of 5-aminolevulinic acid display improved anticancer activity independent and dependent on photoactivation.
J Med Chem, 51(23):7356-7369.


[2] Bolt, E.L., Kryszak, L., Zeilstra-Ryalls, J., 1999. Characterization of the
Rhodobacter sphaeroides 5-aminolaevulinic acid synthase isoenzymes, HemA and HemT, isolated from recombinant
Escherichia coli
.
Eur J Biochem, 265(1):290-299.
[3] Braathen, L.R., Szeimies, R.M., Basset-Seguin, N., 2007. Guidelines on the use of photodynamic therapy for nonmelanoma skin cancer: an international consensus. International Society for Photodynamic Therapy in Dermatology, 2005.
J Am Acad Dermatol, 56(1):125-143.
[4] Burnham, B.F., 1970. δ-Aminolevulinic acid synthase (from
Rhodopseudomonas sphaeroides). Methods in Enzymology, Volume 17, Metabolism of Amino Acids and Amines, Part A. Academic Press,USA :195-200.
[5] Choi, C., Hong, B.S., Sung, H.C., 1999. Optimization of extracellular 5-aminolevulinic acid production from
Escherichia coli transformed with ALA synthase gene of
Bradyrhizobium japonicum
.
Biotechnol Lett, 21(6):551-554.
[6] Choi, H.P., Hong, J.W., Rhee, K.H., 2004. Cloning, expression, and characterization of 5-aminolevulinic acid synthase from
Rhodopseudomonas palustris KUGB306.
FEMS Microbiol Lett, 236(2):175-181.
[7] Choi, H.P., Lee, Y.M., Yun, C.W., 2008. Extracellular 5-aminolevulinic acid production by
Escherichia coli containing the
Rhodopseudomonas palustris KUGB306
hemA gene.
J Microbiol Biotechnol, 18(6):1136-1140.
[8] Fan, L., 2006. Study on Analysis and Stability of 5-Aminolevulinic Acid. MS Thesis, Zhejiang University,Hangzhou, China :
[9] Fu, W.Q., Lin, J.P., Cen, P.L., 2007. 5-Aminolevulinate production with recombinant
Escherichia coli using a rare codon optimizer host strain.
Appl Microbiol Biotechnol, 75(4):777-782.
[10] Fu, W.Q., Lin, H.P., Cen, P.L., 2008. Enhancement of 5-aminolevulinate production with recombinant
Escherichia coli using batch and fed-batch culture system.
Bioresource Technol, 99(11):4864-4870.
[11] Fu, W.Q., Lin, J.P., Cen, P.L., 2010. Expression of a
hemA gene from
Agrobacterium radiobacter in a rare codon optimizing
Escherichia coli for improving 5-aminolevulinate production.
Appl Microbiol Biotechnol, 160(2):456-466.
[12] Kang, D.K., Kim, S.S., Chi, W.J., 2004. Cloning and expression of the
Rhodobacter capsulatus hemA gene in
E. coli for the production of 5-aminolevulinic acid.
J Microbiol Biotechnol, 14(6):1327-1332.
[13] Kiatpapan, P., Murooka, Y., 2001. Construction of an expression vector for propionibacteria and its use in production of 5-aminolevulinic acid by
Propionibacterium freudenreichii
.
Appl Microbiol Biotechnol, 56(1-2):144-149.
[14] Lang, K., Lehmann, P., Bolsen, K., 2001. Aminolevulinic acid: pharmacological profile and clinical indication.
Expert Opin Investig Drugs, 10(6):1139-1156.
[15] Lee, D.H., Jun, W.J., Yoon, J.W., 2004. Process strategies to enhance the production of 5-aminolevulinic acid with recombinant
E. coli
.
J Microbiol Biotechnol, 14(6):1310-1317.
[16] Lin, J.P., Fu, W.Q., Cen, P.L., 2009. Characterization of 5-aminolevulinate synthase from
Agrobacterium radiobacter, screening new inhibitors for 5-aminolevulinate dehydratase from
Escherichia coli and their potential use for high 5-aminolevulinate production.
Bioresource Technol, 100(7):2293-2297.
[17] Liu, X.X., Lin, J.P., Qin, G., 2005. Expression of a new
hemA gene from
Agrobacterium radiobacter in
Escherichia coli for 5-aminolevulinate production.
Chin J Chem Eng, 13(4):522-528.
[18] Pariser, D., Loss, R., Jarratt, M., 2008. Topical methyl-aminolevulinate photodynamic therapy using red light-emitting diode light for treatment of multiple actinic keratoses: a randomized, double-blind, placebo-controlled study.
J Am Acad Dermatol, 59(4):569-576.
[19] Sasaki, K., Watanabe, M., Tanaka, T., 2002. Biosynthesis, biotechnological production and applications of 5-aminolevulinic acid.
Appl Microbiol Biotechnol, 58(1):23-29.
[20] Vanderwerf, M.J., Zeikus, J.G., 1996. 5-Aminolevulinate production by
Escherichia coli containing the
Rhodobacter sphaeroides hemA gene.
Appl Environ Microbiol, 62(10):3560-3566.
[21] Viale, A.A., Wider, E.A., Batlle, A.M.D.C., 1987. Porphyrin biosynthesis in
Rhodopseudomonas palustris—XI. Extraction and characterization of δ-aminolevulinate synthetase.
Comp Biochem Physiol B Comp Biochem, 87(3):607-613.
[22] Wang, J.Q., Wu, J.H., Zhang, Z.M., 2006. Expression of 5-aminolevulinic acid synthase in recombinant
Escherichia coli
.
World J Microbiol Biotechnol, 22(5):461-468.
[23] Warnick, G.R., Burnham, B.F., 1971. Regulation of porphyrin biosynthesis. Purification and characterization of δ-aminolevulinic acid synthase.
J Biol Chem, 246(22):6880-6885.
[24] Xie, L., Hall, D., Eiteman, M.A., 2003. Optimization of recombinant aminolevulinate synthase production in
Escherichia coli using factorial design.
Appl Microbiol Biotechnol, 63(3):267-273.
[25] Xie, L., Eiteman, M.A., Altman, E., 2003. Production of 5-aminolevulinic acid by an
Escherichia coli aminolevulinate dehydratase mutant that overproduces
Rhodobacter sphaeroides aminolevulinate synthase.
Biotechnol Lett, 25(20):1751-1755.
[26] Zaak, D., Sroka, R., Khoder, W., 2008. Photodynamic diagnosis of prostate cancer using 5-aminolevulinic acid: first clinical experiences.
Urology, 72(2):345-348.



Open peer comments: Debate/Discuss/Question/Opinion
<1>