1. Introduction
The fruit is the reproductive organ for higher plants with a complex process of development and senescence, which are regulated by plant hormones (Gillaspy et al.,
1993; Ozga and Reinecke,
2003). Among these hormones, ethylene, the simplest olefin, is involved in many aspects of the plant life cycle, including seed germination, seedling development, leaf expansion, blossoming, senescence, fruit ripening, and responses to biotic and abiotic stresses (Wang et al.,
2002; Lin et al.,
2009). Ethylene is a necessary factor for the initiation of fruit ripening and senescence and promotes the transcription and translation of responsive genes involved in cell-wall metabolism, membrane metabolism, and fruit softening by activating ethylene signaling transduction (Wang et al.,
2002; Alonso and Stepanova,
2004).
The biosynthesis of ethylene begins with the production of
S-adenosylmethionine (SAM), which is catalyzed by SAM synthetase (EC 2.5.1.6) from methionine. SAM is converted to 1-aminocyclopropane-1-carboxylic acid (ACC) and 5-methylthioadenosine (MTA) by ACC synthase (ACS; EC 4.4.1.14). ACC is subsequently oxidized by ACC oxidase (ACO; EC 1.14.17.4) to yield ethylene and CO
2 (Yang and Hoffmann,
1984; Kende,
1993). ACS and ACO are the key enzymes producing ethylene, are encoded by a multigene family, and have been isolated from numerous plant species.
ACS and
ACO genes show different expression profiles in plant tissues, and respond to different stimuli, including low oxygen, mechanical damage, high temperature, and hormones (Arteca and Arteca,
1999; Rieu et al.,
2005; Choudhury et al.,
2008).
Ethylene is one of the most important factors regulating softening, senescence, and abscission of fruit, and the concentration of ethylene increases gradually during the development of climacteric fruit. ACS and ACO act as accelerators in fruit ripening by catalyzing the production of ethylene at the late fruit ripening stage, leading to irreversible senescence (Chen et al.,
2003; Tatsuki et al.,
2006; Cara and Giovannoni,
2008; Varanasi et al.,
2011). It has been shown that once the expression levels of
ACS and
ACO genes were suppressed or mutated, the production of ethylene was down-regulated and the shelf life of fruit was also prolonged. Oeller et al. (
1991) transformed the antisense messenger RNA (mRNA) of the
ACS gene in tomato and inhibited fruit ripening. Ayub et al. (
1996) generated transgenic melons by introducing an antisense
ACO gene, and ethylene production of transgenic fruit was down-regulated compared with untransformed fruit. The silencing of
LeACS2 by vacuum-infiltration and the tobacco rattle virus (TRV)-based virus-induced gene silencing (VIGS) method also leads to a significant delay in the post-harvest ripening and senescence of tomato fruit (Xie et al.,
2006). Moreover, null mutations of the
MdACS1 and
MdACS3 genes have shown no or very low expression levels of ripening-related genes and maintained firmness in apple fruit (
Malus domestica) (Sunako et al.,
1999; Wang et al.,
2009).
Mulberry (
Morus alba L.) is a deciduous tree and an economically important food crop for the domesticated silkworm (
Bombyx mori L.). Mulberry has multiple uses in ecology, pharmaceuticals, and traditional Chinese medicine (He et al.,
2013). Its bark is also used in paper production. In addition, mulberry fruit is one of the most popular fruits worldwide and is especially appreciated for its unique flavor in China. However, the commercialization of mulberry fruit is limited by its short maturity stages and shelf-life. The effective strategy to make use of mulberry fruit is to strengthen deep processing or to utilize ethylene inhibitors, such as silver thiosulfate, 1-methylcyclopropene (1-MCP), aminoethoxyvinylglycine, and 2,5-norbornadiene, which delay ripening and senescence (Blankenship and Dole,
2003). Furthermore, a potential tool to delay the ripening of mulberry fruit and to prolong its shelf-life is the use of transgenic approaches to down-regulate the expression of ethylene biosynthesis-related genes.
The ethylene biosynthesis pathway has been detected in some plant species, like
Arabidopsis thaliana, tomato (
Lycopersicon esculentum), and apple. However, there is little knowledge on ethylene biosynthesis in mulberry. In this study, we report the isolation and characterization of five
ACS genes and two
ACO genes from mulberry, and demonstrate their expression levels in different organs and fruit with different development stages following treatment with abscisic acid (ABA) and ethephon. We hope this work will provide insights into the developmental functions of these genes and lay a foundation for further understanding the mechanism of mulberry fruit development and ripening.
2. Materials and methods
2.1. Plant materials and treatments
Different tissues (e.g., root, stem, stem epidermis, petiole, leaf, and fruit) were collected from the mulberry cultivar “Changjiang 1”, which is grown in the mulberry garden of Southwest University, Chongqing, China. The mulberry fruit were sampled after full-bloom stages of 10, 20, 29, 36, and 40 d. All plant materials were frozen in liquid nitrogen and stored at −80 °C.
The immature fruit on 20 d after full-bloom (DAF) with a partial red color were collected from trees treated with 264 mg/L ABA (containing 0.1% Tween-20) and 100 mg/L ethephon (containing 0.1% Tween-20) for 5 min at 25 °C; the control fruit were dipped in double-distilled water (ddH
2O) (Ren and Leng,
2010). After dipping, the fruit were kept at 25 °C for 5 d and were immediately frozen in liquid nitrogen and stored at −80 °C.
2.2. Isolation of RNA and synthesis of the first strands of complementary DNA (cDNA)
Total RNA was extracted from root, stem, stem epidermis, petiole, and leaf using the RNA Extraction Kit (TaKaRa, Japan) as described in the manufacturer’s instructions. Total RNA of fruit was extracted using the RNA Extraction Kit Transzol Plant (TransGen Biotech, China). To remove genomic DNA, the RNA samples were digested by DNase I (TaKaRa, Japan). The cDNA was synthesized from 3 μg of DNA-free RNA with the reverse transcriptional Moloney murine leukaemia virus (M-MLV; Promega) following the manufacturer’s protocol. Ten-fold diluted cDNA was used in quantitative reverse transcriptase polymerase chain reaction (qRT-PCR).
2.3. Identification and isolation of ACS and ACO
ACS and
ACO genes from other plant species were downloaded from GenBank (
http://www.ncbi.nlm.nih.gov/genbank/), and were used as queries in BLAST searches against Morus Genome Database (
http://morus.swu.edu.cn/morusdb/). This yielded non-redundant full mRNA sequences of
ACS and
ACO genes. The candidate genes were identified using BLASTN and SMART (
http://smart.embl-heidelberg.de/). Five
ACS genes and two
ACO genes were finally selected for further research.
2.4. Sequence alignment and phylogenetic analysis
The similarities of
ACS and
ACO genes were analyzed by BLASTN. About 64 ACS and 46 ACO amino acid sequences were downloaded from the National Center of Biotechnology Information (NCBI) and used for alignment with putative protein sequences of mulberry
ACS and
ACO genes given by MUSCLE 3.6, and a phylogenetic tree was constructed by MEGA 5.0 software.
2.5. qRT-PCR expression analysis of mulberry ACS and ACO genes
The primers used for qRT-PCR were designed by the online tool of the GeneScript Company of China (
http://www.genscript.com.cn/index.html) (Table
1). For the analyses of
ACS and
ACO gene expression, 20 μl total reaction volume composed of 10 μl of 2× SYBR® Premix Ex Taq II (TaKaRa, Japan), 0.8 μl of each primer (10 μmol/L), 0.4 μl of 50× ROX Reference Dye II (TaKaRa, Japan), 2 μl of cDNA, and 6 μl of ddH
2O. The thermal parameters for qRT-PCR were: 95 °C for 30 s, followed by 40 cycles of 94 °C for 5 s and 60 °C for 30 s, with a final extension at 72 °C for 10 min. The
MaACTIN3 (HQ163775.1) gene was used as an internal control. Relative expression levels were calculated using the fruit after full-bloom stage of 10 d set at 1.
Table 1
Primers for qRT-PCR
| Gene |
Primer (5′ to 3′) |
|
MaACS1
|
Forward: GCAACACTGACCTCATCCAC |
|
Reverse: TGCGAGGATACTAATCCGAA |
|
MaACS2
|
Forward: GTCAACGAGGTCAAGCTCAA |
|
Reverse: GTTTACGGTCTCCTCCTCCA |
|
MaACS3
|
Forward: TGGCTTCCAAATCACTGAAG |
|
Reverse: GGTTGTGTTCGTTCGATGTC |
|
MaACS4
|
Forward: CTCAAACGGCTTCAGAATCA |
|
Reverse: AGCTCGTCAGGAGTCAAGGT |
|
MaACS5
|
Forward: CAGCTTCTGTTGAGCTCTCG |
|
Reverse: CATCATAAGCTTTCCAGCCA |
|
MaACO1
|
Forward: AAGGTGATGAGGGAATTTGC |
|
Reverse: GGTCCCTTTGAGCCATAGAA |
|
MaACO2
|
Forward: TCTTGGACTGGAGAAAGGGT |
|
Reverse: CCCTTGATCAGGTCTGGTTT |
|
MaACTIN3
|
Forward: GCATGAAGATCAAGGTGGTG |
|
Reverse: CATCTGCTGGAAGGTGCTAA |
3. Results
3.1. Identification and isolation of MaACS and MaACO genes
ACS and
ACO gene sequences from other plants were used to search the mulberry genome sequences and the obtained mRNA sequences were screened by the BLASTN program; their encoded amino acids were analyzed by the SMART program. Five
ACS genes and two
ACO genes were finally screened, which were named as
MaACS1,
MaACS2,
MaACS3,
MaACS4,
MaACS5,
MaACO1, and
MaACO2.
MaACO1 had previously been submitted to the NCBI (GenBank Accession No. KC709482).
MaACO1 may be associated with tissue aging or senescence and respond to stress in mulberry (Pan and Lou,
2008).
The full-length genomic sequences of
MaACS genes ranged from 1660 bp (
MaACS5) to 2732 bp (
MaACS2) and the open reading frame (ORF) length ranged from 1341 bp (
MaACS5) to 1491 bp (
MaACS2), encoding polypeptides ranging from 446 to 496 amino acids with predicted molecular masses ranging from 50.16 to 55.57 kDa. The full-length genomic sequences of
MaACO genes were 1666 bp (
MaACO1) and 1526 bp (
MaACO2) and the ORF lengths were 960 and 969 bp, encoding polypeptides of 319 and 322 amino acids with predicted molecular masses of 36.38 and 36.48 kDa, respectively (Table
2).
Table 2
Gene analyses of MaACS and MaACO
| Gene |
Genomic full-length (bp) |
ORF length (bp) |
Amino acids |
MW (kDa) |
pI |
Gene ID |
|
MaACS1
|
2196 |
1461 |
486 |
54.68 |
6.47 |
Morus024218 |
|
MaACS2
|
2732 |
1491 |
496 |
55.57 |
6.77 |
Morus012919 |
|
MaACS3
|
1782 |
1416 |
471 |
53.42 |
8.82 |
Morus007775 |
|
MaACS4
|
2044 |
1404 |
467 |
52.50 |
5.93 |
Morus007092 |
|
MaACS5
|
1660 |
1341 |
446 |
50.16 |
5.55 |
Morus027243 |
|
MaACO1
|
1666 |
960 |
319 |
36.38 |
5.35 |
Morus004820 |
|
MaACO2
|
1526 |
969 |
322 |
36.48 |
5.22 |
Morus014137 |
MW: molecular weight of the MaACS and MaACO proteins; pI: isoelectric point of the MaACS and MaACO proteins; ID: IDs of MaACS and MaACO in the Morus Genome Database
3.2. Structure of MaACS and MaACO genes
To reveal the intron and exon organizations of
MaACS and
MaACO genes, we compared all obtained ORF sequences with the corresponding genomic DNA sequences (Fig.
1). The
MaACS sequences could be divided into two groups. The first group included
MaACS1,
MaACS2,
MaACS3, and
MaACS4, which have a 4-exon/3-intron structure, and
MaACS5 has a 3-exon/2-intron structure. Both
MaACO1 and
MaACO2 have a 4-exon/3-intron structure.
Fig.1
Schematic diagrams of MaACS and MaACO genes with exons (black boxes) and introns (lines between exons)
3.3. Multiple-sequence alignments and phylogenetic analysis of MaACS genes
Sequence similarity matrix analysis of
MaACS amino acids has revealed that the sequence identity ranged from 50% to 69% (Table
3). The sequence identities were higher between
MaACS1 and
MaACS2 (69%) and between
MaACS3 and
MaACS4 (68%).
Table 3
Sequence similarity matrix of MaACS
| Gene |
Sequence identity (%)
|
|
MaACS1
|
MaACS2
|
MaACS3
|
MaACS4
|
MaACS5
|
|
MaACS1
|
100 |
|
MaACS2
|
69 |
100 |
|
MaACS3
|
53 |
56 |
100 |
|
MaACS4
|
50 |
54 |
68 |
100 |
|
MaACS5
|
57 |
60 |
60 |
58 |
100 |
The amino acids encoded by
MaACS genes were aligned by the Clustal W program and have the pyridoxal phosphate (PLP)-dependent enzyme domain, which is a co-factor associated with ACS catalyzing the conversion of
S-adenosyl-L-methionine (AdoMet) to ACC, predicted by the SMART program. It was shown that ACS proteins could be divided into three main types differing in the C-terminal sequences. Type I proteins have an extended C-terminus containing three conserved serine residues that are targets for phosphorylation by mitogen-activated protein kinase 6 (MPK6) and a conserved serine residue that is a phosphorylation site for calcium-dependent protein kinase (CDPK) (Tatsuki and Mori,
2001; Chae et al.,
2003; Liu and Zhang,
2004; Sebastià et al.,
2004). Type II proteins have a shorter C-terminus that has only the CDPK phosphorylation site. Type III proteins have a very short C-terminal extension that lacks both phosphorylation sites. The result of multiple-sequence alignment against MaACS proteins showed that MaACS1 and MaACS2 were type I, MaACS3 and MaACS4 were type II, and MaACS5 was type III (Fig.
2).
Fig.2
Amino acid alignment of the inferred MaACS gene products
The sequence in the red box indicates the PLP-dependent enzyme domain. Black boxes and black circles (●) indicate CDPK and MPK6 active sites, respectively. ▼ indicates the active sites associated with heterodimeric interactions (Note: for interpretation of the references to color in this figure legend, the reader is referred to the web version of this article)
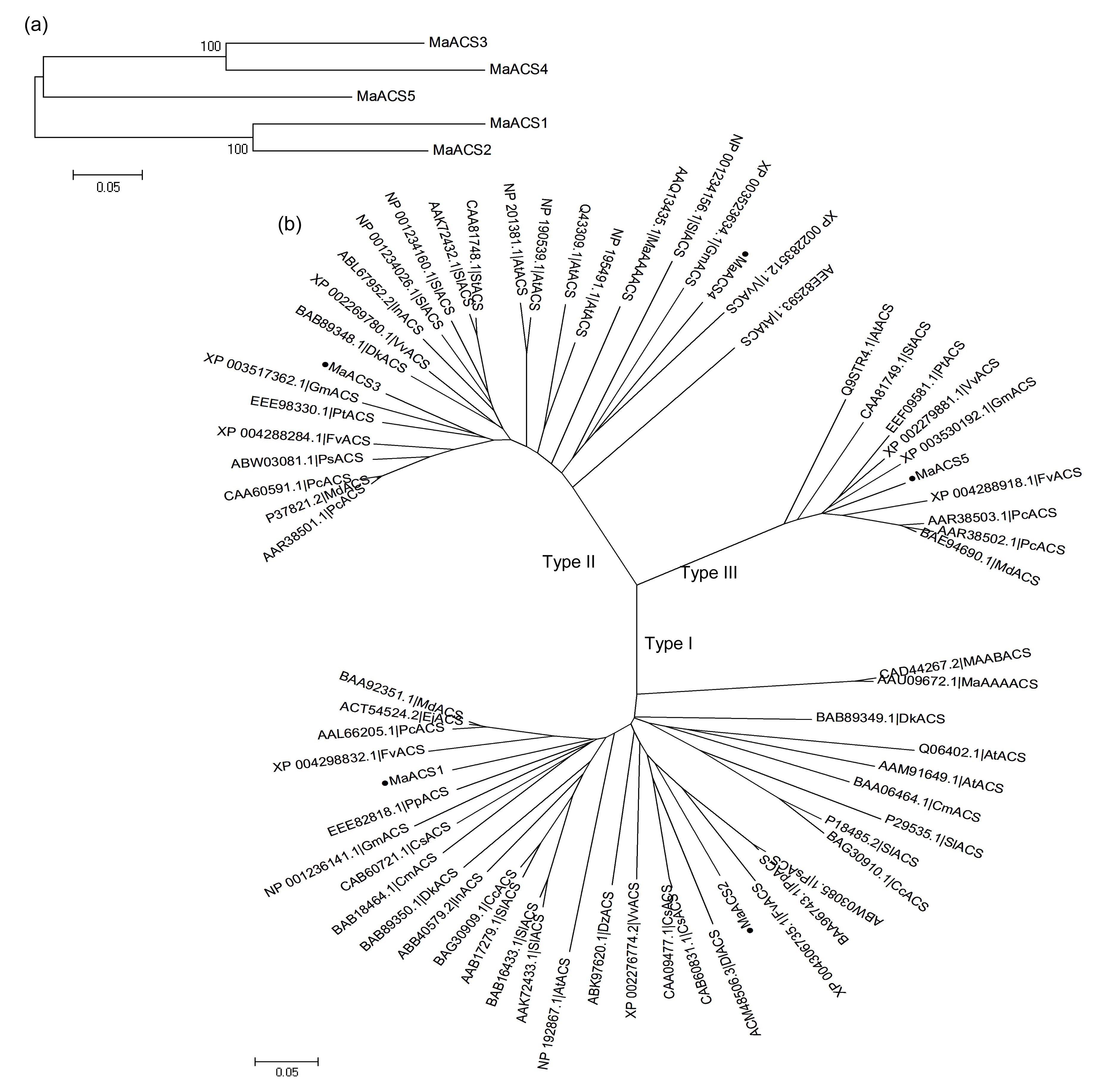
The amino acids of MaACS were aligned by the Muscle 3.6 program and a phylogenetic tree was constructed by MEGA 5.0. The resulting tree showed that MaACS1 and MaACS2, MaACS3 and MaACS4, and MaACS5 were three branches, respectively (Fig.
3a). A phylogenetic tree was also constructed for ACS from mulberry with other plants via the neighbor-joining method (Fig.
3b). The tree was divided into three groups: I, II, and III; MaACS1 and MaACS2 were group I, MaACS3 and MaACS4 were group II, and MaACS5 was group III. This result is consistent with the classification based on the difference in the C-terminal sequences of MaACS.
Fig.3
Phylogenetic tree analysis of MaACS proteins
(a) Phylogenetic relationships between MaACSs; (b) Phylogenetic relationships between MaACS proteins and other ACS proteins from other plant species
ACS proteins can act as homodynamic or heterodimeric proteins to form active heterodimers enhancing the isozyme diversity of the
ACS gene family and providing physiological multifunctionality by operating in different SAM concentrations in various cells and tissues during plant growth and development (Tsuchisaka and Theologis,
2004). In this study, Y92 and K278 residues of MaACS proteins are predicted as the active sites associated with heterodimeric interactions (Fig.
2).
3.4. Multiple-sequence alignments and phylogenetic analysis of MaACO genes
ACO is a member of a superfamily of non-haem iron-dependent oxygenases, most of which utilize Fe(II) as a co-factor and 2-oxoglutarate (2OG) as a co-substrate (John,
1991; Bidonde et al.,
1998). The domain of
MaACO encoding protein sequences was analyzed by the SMART program and NCBI. The results revealed that the sequences of amino acids ranging from 156 to 256 have the 2OG-Fe(II) oxygenase domain (Fig.
4), which indicates that ACO is a member of the ketoglutaric acid dioxygenase family. Crystal structure studies of the
Petunia hybrida ACO showed that it forms a complex with Fe(II) and Cu(II) and contains a single Fe(II) atom binding site coordinated by three residues (His177, Asp179, and His234), and Arg175 and Arg244 are proposed to be involved in binding bicarbonate; this leads to the activation of the ACO enzyme (Zhang et al.,
2004). Here, as reported previously, H177, D179, and H234 in MaACO1 and MaACO2 proteins were predicted and identified as Fe(II) binding sites, and R175 and R244 were predicted as 2OG binding sites (Fig.
4).
Fig.4
MaACO protein sequence alignments with other ACO proteins
The sequence in the box indicates the 2OG-Fe(II) oxygenase enzyme domain. ▼ indicates Fe(II) binding sites and ● indicates the 2OG binding sites
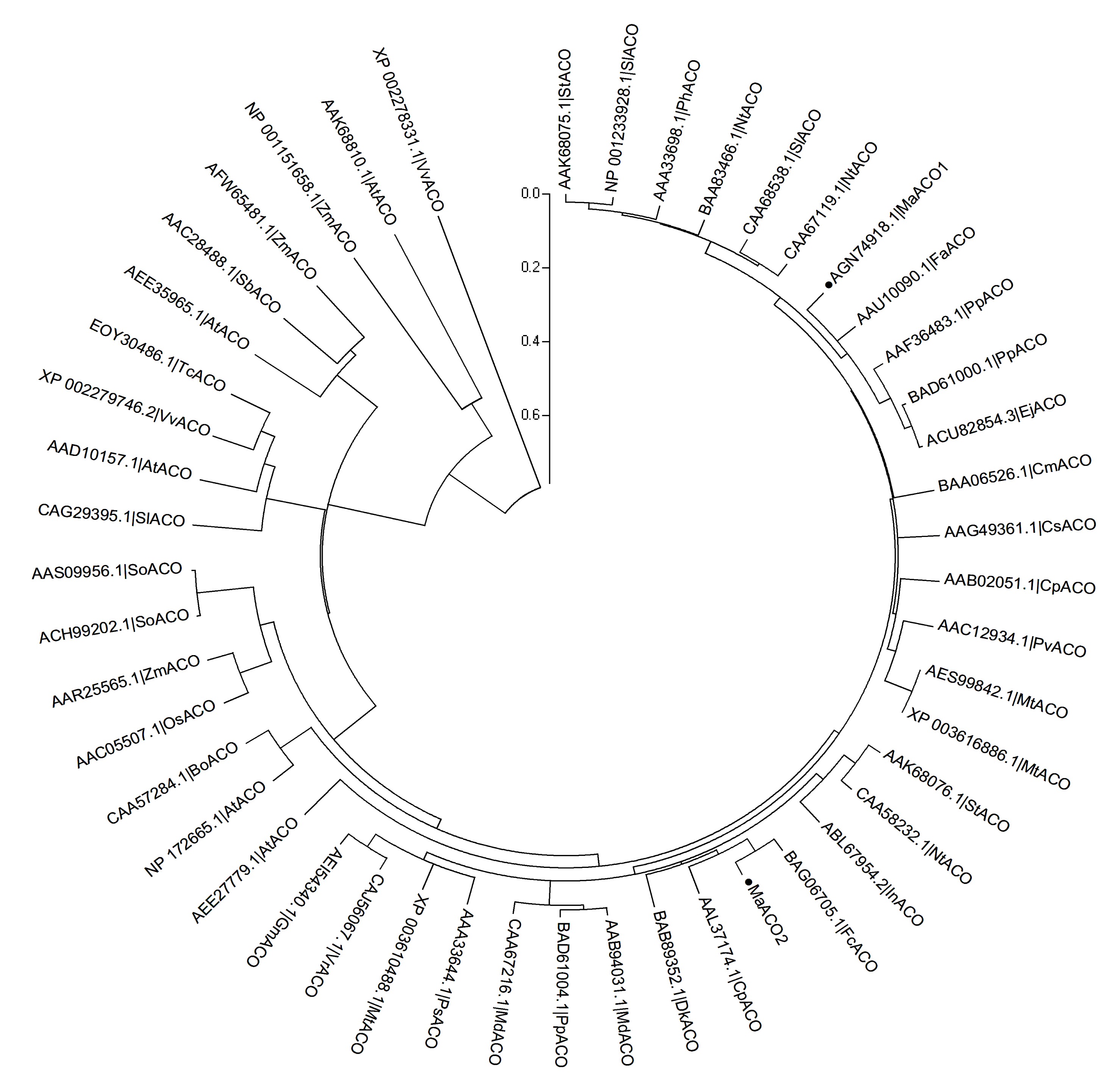
Multiple-sequence alignment results showed that ACO amino acids are relatively conserved with high similarities in higher plants. The results of phylogenetic analysis of MaACO suggested that VvACO (XP_002278331.1), AtACO (AAK68810.1), and ZmACO (NP_001151658.1) could be clustered into the same clades, and MaACO1 and MaACO2 could be clustered into a branch with another 43 ACO proteins from NCBI (Fig.
5).
Fig.5
Phylogenetic relationships between MaACO proteins and other ACO proteins from other species
3.5. Tissue specificity in qRT-PCR expression analyses of MaACS and MaACO genes
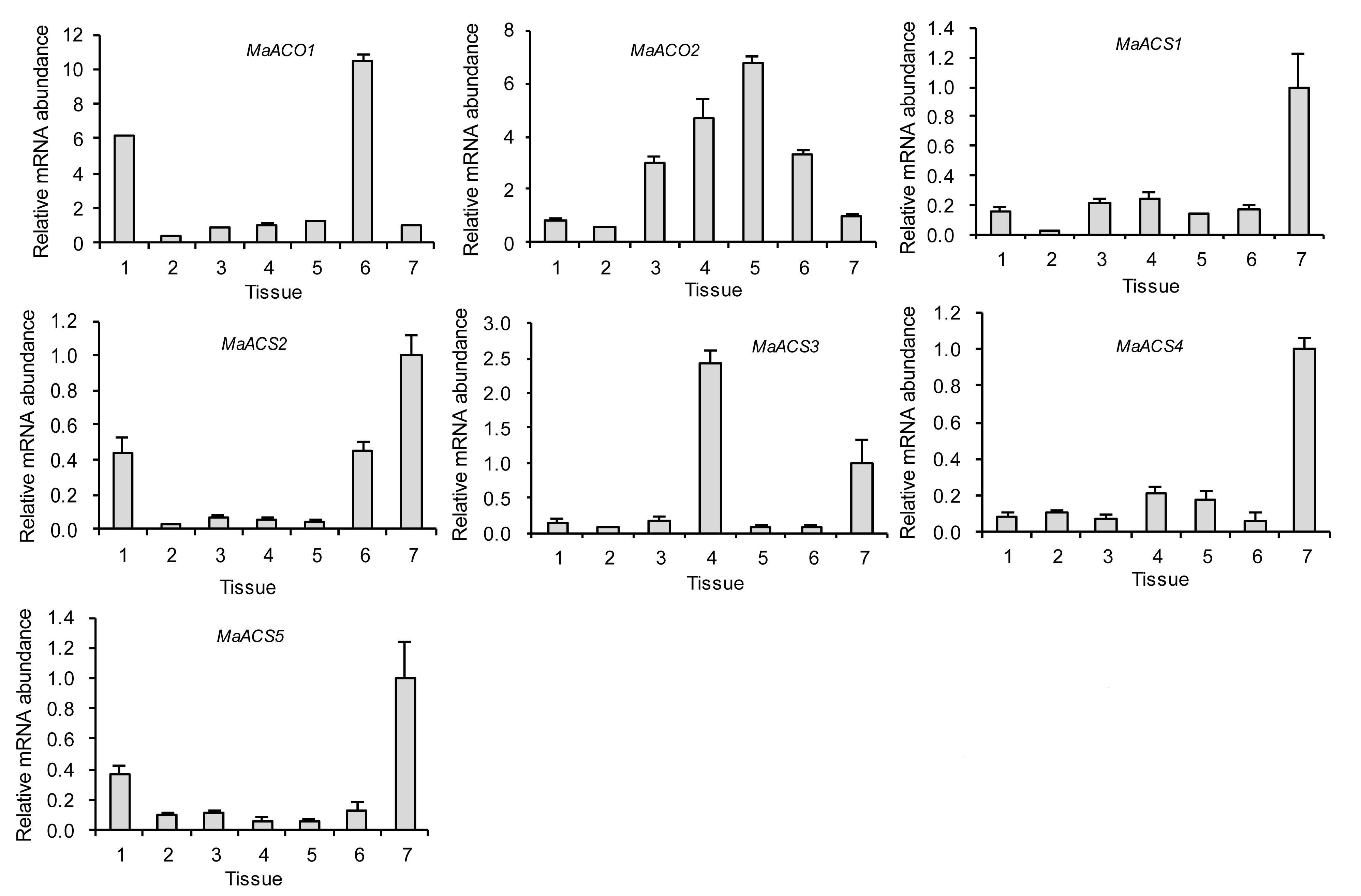
Tissue-specific expression analysis revealed that
MaACS and
MaACO genes were differentially expressed in various mulberry tissues (Fig.
6). Within the
MaACS gene family,
MaACS1 and
MaACS4 showed similar expression patterns, with transcripts strongly expressed in fruit and more weakly in roots, stem, stem epidermis, petioles, and leaves.
MaACS2 and
MaACS5 showed somewhat similar expression patterns, and had higher expression in roots, old leaves, and fruit, and lesser levels in others, although
MaACS5 transcripts were also low in roots.
MaACS3 transcripts were more abundant in petiole and fruit tissues, and the expression level was higher in petioles than in fruit.
Fig.6
Expression of MaACO and MaACS genes in various mulberry tissues
Tissue: 1, root; 2, stem; 3, stem epidermis; 4, petiole; 5, immature leaves; 6, mature leaves; 7, fruitlet on 10 DAF. Columns heights indicate relative mRNA abundance, and the abundance of fruitlet on 10 DAF (Tissue 7) was set to 1. Error bars on each column indicate standard errors (SEs) from three replications
The expression of
MaACO1 and
MaACO2 showed different patterns. The transcript of
MaACO1 had higher levels in roots and old leaves, and lower levels in other tissues.
MaACO2 had higher levels in stem epidermis, petioles, and immature and old leaves, and lower levels in other tissues.
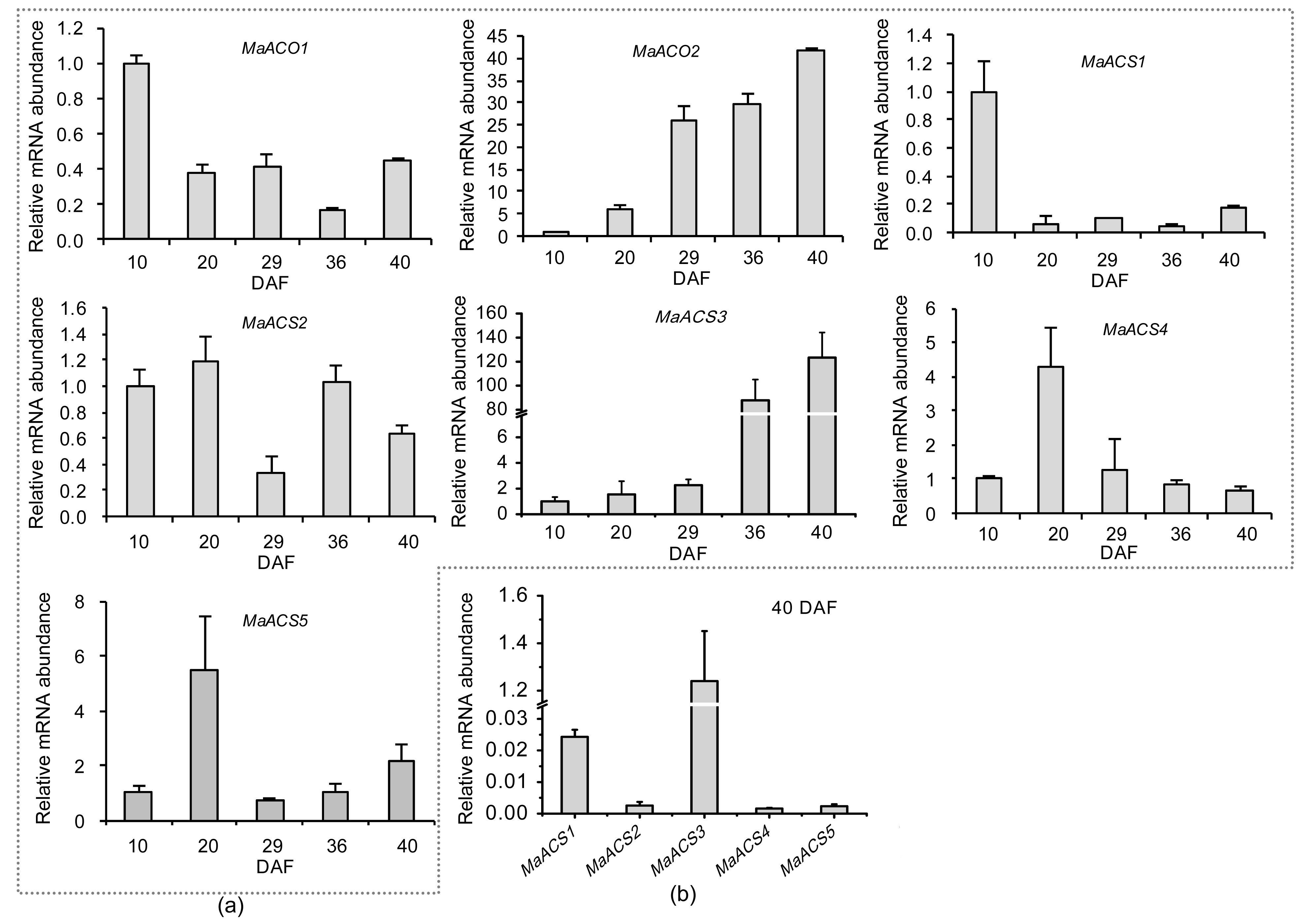
3.6. qRT-PCR expression analysis of MaACS and MaACO genes in fruit
The expression of the different
MaACS and
MaACO genes showed two patterns during the development of mulberry fruit (Fig.
7).
MaACS1,
MaACS2,
MaACS4,
MaACS5, and
MaACO1 had strong peaks of expression in early stages of fruit, and decreased throughout development. However, transcript abundance of
MaACS3 and
MaACO2, unlike other genes, increased rapidly throughout fruit development.
Fig.7
Expression of MaACO and MaACS genes during mulberry fruit development (a) and the expression levels of MaACS genes in the fruit of 40 DAF (b)
Each column height indicates relative mRNA abundance, and the abundance of fruitlet on 10 DAF was set to 1. Error bars on each column indicate standard errors (SEs) from three replications
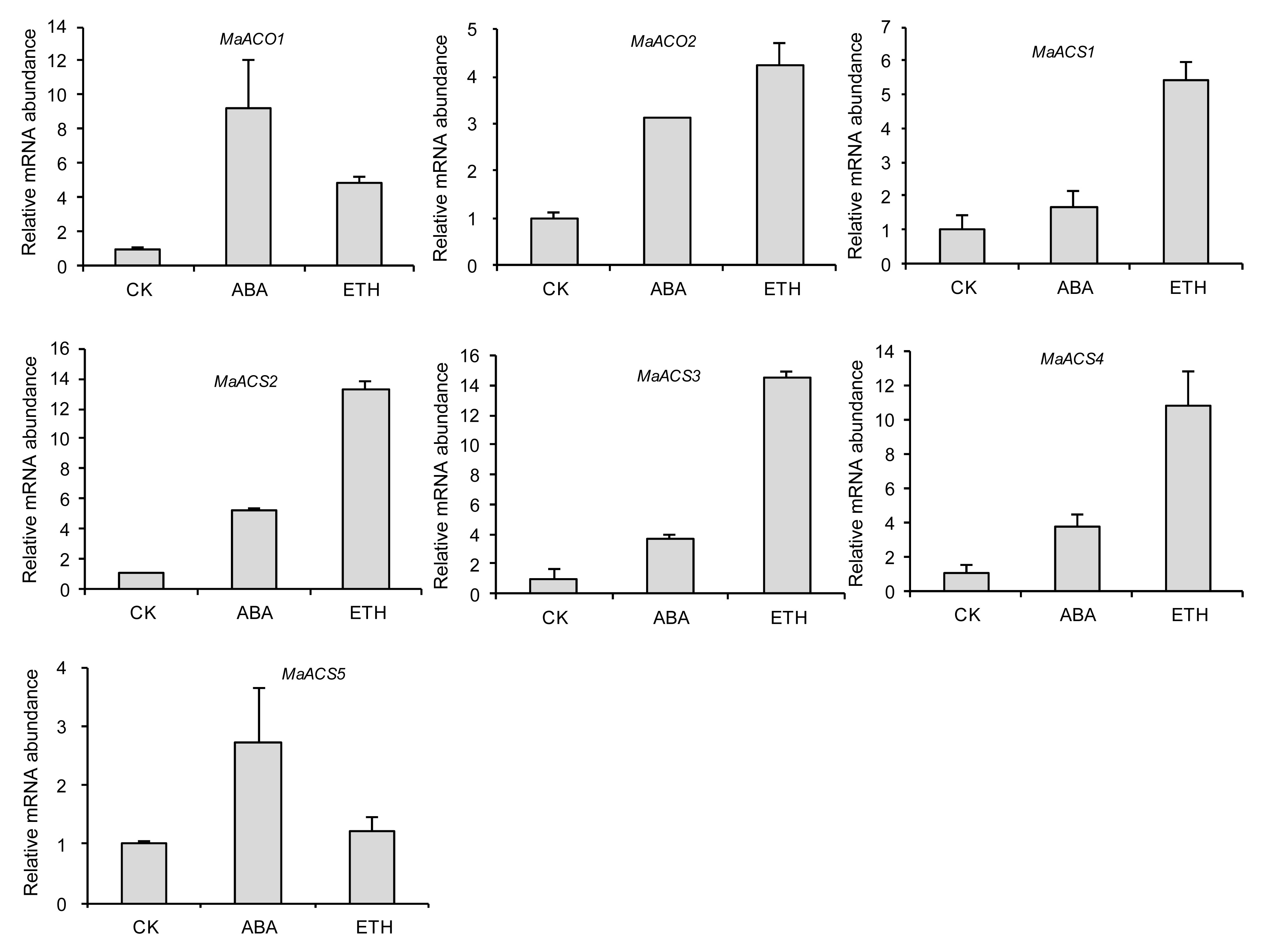
The mulberry fruit on 20 DAF were treated with ABA and ethephon, and fruit turned the full red color (data not shown). Transcript abundance of
MaACS and
MaACO genes were up-regulated by ABA and ethephon (Fig.
8). This indicated that ABA and ethephon could regulate the transcription of ethylene biosynthesis-related genes.
Fig.8
Expression of MaACO and MaACS genes in fruit incubated with ddH2O (CK), ABA (264 mg/L), and ethephon (ETH, 100 mg/L)
Columns heights indicate relative mRNA abundance, and the abundance of fruit treated ddH2O was set to 1. Error bars on each column indicate standard errors (SEs) from three replications
4. Discussion
ACS and ACO are the key enzymes producing ethylene, and are typically encoded by multiple
ACS and
ACO genes, and some members of these gene families are differentially regulated to respond to different cues in various tissues throughout plant development. In this study, we have isolated and analyzed five
ACS genes and two
ACO genes from mulberry genome database. Sequence alignments and phylogenetic analyses of
MaACO1 and
MaACO2 showed that sequence identity is 82% and their amino acids are conserved compared with ACO proteins from other species. Multiple-sequence alignments and phylogenetic analyses of
MaACS genes show that
ACS genes have the PLP-dependent enzyme domain and could be divided into three types (I, II, and III) in higher plants based on the difference in the C-terminal sequences. Like
Arabidopsis thaliana, tomato, apple, and grape (
Vitis vinifera),
ACS is classified into three types in mulberry:
MaACS1 and
MaACS2 are type I,
MaACS3 and
MaACS4 are type II, and
MaACS5 is type III.
Fruit can be classified into two groups, climacteric and non-climacteric fruit, based on pattern of respiration and ethylene release during maturation and ripening. Further, the ethylene biosynthesis in fruit is controlled by two systems (McMurchie et al.,
1972): system I is responsible for ethylene production during normal vegetative growth in all tissues including climacteric and non-climacteric fruit; system II only exists in climacteric fruit, and is essential for the burst of ethylene production during the ripening of fruit (Seymour et al.,
1993). The transition from system I to system II is considered to be an important step in fruit ripening (Lelièvre et al.,
1997; Cara and Giovannoni,
2008). In climacteric fruit, system I-like
ACS and
ACO genes showed lower expression levels in the later stages of fruit, but transcript abundance of system II-like genes increased rapidly throughout fruit development. The expression of the different
MaACS and
MaACO genes showed two patterns during the development of mulberry fruit.
MaACS1,
MaACS2,
MaACS4,
MaACS5, and
MaACO1 had strong peaks of expression in early stages of fruit, and decreased throughout development. However, transcript abundance of
MaACS3 and
MaACO2, unlike other
MaACS and
MaACO genes, increased rapidly throughout fruit development. However, these data could not support the view that
MaACS3 and
MaACO2 are responsible for system II ethylene biosynthesis in ripening fruits and that other genes are responsible for system I, because all
MaACS and
MaACO genes were stimulated by ethephon as shown in Fig.
8; more studies are needed to support
MaACS and
MaACO gene classification.
Within the
MaACS genes, the expression pattern of
MaACS3 was different from those of other
MaACS genes, and its transcript abundance was increased gradually during fruit development, especially increased by 38.8-fold from 29 to 36 DAF of fruit. In addition, the expression level of
MaACS3 was higher than those of the other four
MaACS genes in the fruit on 40 DAF. Therefore, the high accumulation of
MaACS3 is closely associated with the production of ethylene in later stages of mulberry fruit. It is possible that mulberry fruit is climacteric on the basis of the results from Luo (
2003a;
2003b), and mulberry fruit certainly decays readily. It is probable that using
MaACS3 as the target gene to suppress its expression in the later stages of mulberry fruit development will prolong shelf-life.
Tissue-specific expression analysis revealed that the
MaACS and
MaACO genes were differentially expressed in various mulberry tissues. Within the
MaACS gene family, the transcript abundance in the fruit on 10 DAF was much higher than that in other tissues. However, the expression of
MaACO genes showed different patterns, the transcriptional levels of
MaACO1 and
MaACO2 in fruit being equal to or lower than those in other tissues. It is indicated that
MaACS genes, unlike
MaACO genes, tend to be expressed in fruit compared with other tissues. Promoters are the key factor driving time and space expression of the correspondent gene, and fruit-specific promoters can control gene expression in fruit but not in other tissues. There are some promoters, like tomato E8/E4 promoter, peach (
Prunus persica)
ACO1 promoter, and apple
ACO1 promoter, being expressed specifically in fruit (Deikman et al.,
1992; Blume and Grierson,
1997; Moon and Callahan,
2004), and the target genes fused to these promoters could overexpress specifically in fruit. There are no reports on fruit-specific promoters using the promoter of
ACS genes. Based on the expression analysis of
MaACS genes in different tissues and fruit at different stages,
MaACS genes are potential targets to develop fruit-specific promoters. Considering that
MaACS1,
MaACS2,
MaACS4, and
MaACS5 have lower expression levels in the later stages of fruit (Fig.
7b), it is probable that we could use
MaACS3 promoter as a candidate to research the fruit-specific promoter in mulberry.
ABA and ethylene are considered as the key factors regulating fruit maturity and senescence. Some research has confirmed that ABA plays an important role in triggering ethylene biosynthesis and softening during the ripening of fruit (Zhang et al.,
2009; Zaharah et al.,
2013). ABA can stimulate the production of ethylene by promoting the activities of ethylene biosynthesis enzymes ACS and ACO, and regulating the expression of its encoded genes, thus starting the ripening process. ABA, like ethephon, can rapidly up-regulate the expression levels of
MaACS and
MaACO genes. This indicates that ABA can promote the biosynthesis of ethylene in mulberry fruit by enhancing the expression of
ACS and
ACO genes. Sun et al. (
2010) thought that the productions of ABA and ethylene at the onset of berry ripening and after harvest are affected by each other. Thus, the relationship between these two hormones and how they affect fruit ripening need further studies.
5. Conclusions
We are the first to report
ACS and
ACO genes from mulberry and demonstrate their expression profiles. Tissue-specific expression analysis revealed that
MaACS and
MaACO genes showed different patterns in various mulberry tissues and two patterns throughout the development of mulberry fruit.
MaACS genes also tended to be expressed in fruit rather than in other tissues. Further, the transcription levels of
MaACS and
MaACO genes could be up-regulated by ABA and ethephon. In future practice, we may be able to use
MaACS3 as the target gene to develop a fruit-specific promoter and to prolong the shelf-life of mulberry fruit by down-regulating its expression.
* Project supported by the National Natural Science Foundation of China (No. 31360190), the Special Fund for Agro-scientific Research in the Public Interest of China (No. 201403064), and the China Agriculture Research System (No. CARS-22-ZJ0102) Chang-ying LIU, Rui-hua LÜ, Jun LI, Ai-chun ZHAO, Xi-ling WANG, Umuhoza DIANE, Xiao-hong WANG, Chuan-hong WANG, Ya-sheng YU, Shu-mei HAN, Cheng LU, and Mao-de YU declare that they have no conflict of interest.References
[1] Alonso, J.M., Stepanova, A.N., 2004. The ethylene signaling pathway.
Science, 306(5701):1513-1515.


[2] Arteca, J.M., Arteca, R.N., 1999. A multi-responsive gene encoding 1-aminocyclopropane-1-carboxylate synthase (
ACS6) in mature
Arabidopsis leaves.
Plant Mol Biol, 39(2):209-219.
[3] Ayub, R., Guis, M., Ben Amor, M., 1996. Expression of ACC oxidase antisense gene inhibits ripening of cantaloupe melon fruits.
Nat Bitechnol, 14(7):862-866.
[4] Bidonde, S., Ferrer, M.A., Zegzouti, H., 1998. Expression and characterization of three tomato 1-aminocyclopropane-1-carboxylate oxidase cDNAs in yeast.
Eur J Biochem, 253(1):20-26.
[5] Blankenship, S.M., Dole, J.M., 2003. 1-Methylcyclopropene: a review.
Posth Biol Technol, 28(1):1-25.
[6] Blume, B., Grierson, D., 1997. Expression of ACC oxidase promoter-GUS fusions in tomato and
Nicotiana plumbaginifolia regulated by developmental and environmental stimuli.
Plant J, 12(4):731-746.
[7] Cara, B., Giovannoni, J.J., 2008. Molecular biology of ethylene during tomato fruit development and maturation.
Plant Sci, 175(1-2):106-113.
[8] Chae, H.S., Faure, F., Kieber, J.J., 2003. The
eto1,
eto2, and
eto3 mutations and cytokinin treatment increase ethylene biosynthesis in
Arabidopsis by increasing the stability of ACS protein.
Plant Cell, 15(2):545-559.
[9] Chen, Y.T., Lee, Y.R., Yang, C.Y., 2003. A novel papaya ACC oxidase gene (
CP-ACO2) associated with late stage fruit ripening and leaf senescence.
Plant Sci, 164(4):531-540.
[10] Choudhury, S.R., Roy, S., Sengupta, D.N., 2008. Characterization of transcriptional profiles of
MA-ACS1 and
MA-ACO1 genes in response to ethylene, auxin, wounding, cold and different photoperiods during ripening in banana fruit.
J Plant Physiol, 165(18):1865-1878.
[11] Deikman, J., Kline, R., Fischer, R.L., 1992. Organization of ripening and ethylene regulatory Regions in a fruit-specific promoter from tomato (
Lycopersicon esculentum).
Plant Physiol, 100(4):2013-2017.
[12] Gillaspy, G., Ben-David, H., Gruissem, W., 1993. Fruits: a developmental perspective.
Plant Cell, 5(10):1439-1451.
[13] He, N., Zhang, C., Qi, X., 2013. Draft genome sequence of the mulberry tree
Morus notabilis
.
Nat Commun, 2:2455
[14] John, P., 1991. How plant molecular biologists revealed a surprising relationship between two enzymes, which took an enzyme out of a membrane, where it was not located and put it into the soluble phase where it could be studied.
Plant Mol Biol Rep, 9(3):192-194.
[15] Kende, H., 1993. Ethylene biosynthesis.
Annu Rev Plant Physiol Plant Mol Biol, 44(1):283-307.
[16] Lelivre, J.M., Latch, A., Jones, B., 1997. Ethylene and fruit ripening.
Physiol Plant, 101(4):727-739.
[17] Lin, Z., Zhong, S., Grierson, D., 2009. Recent advances in ethylene research.
J Exp Bot, 60(12):3311-3336.
[18] Liu, Y., Zhang, S., 2004. Phosphorylation of 1-aminocyclopropane-1-carboxylic acid synthase by MPK6, a stress-responsive mitogen-activated protein kinase, induces ethylene biosynthesis in
Arabidopsis
.
Plant Cell, 16(12):3386-3399.
[19] Luo, Z., 2003. Effect of modified atmosphere storage on the cell wall component and cell wall hydrolase activity of mulberry fruit.
J Fruit Sci, (in Chinese),20(3):214-217.
[20] Luo, Z., 2003. Effect of modified atmosphere storage on the physiology of mulberry fruit.
J Chin Inst Food Sci Technol, (in Chinese),3(3):51-54.
[21] McMurchie, E.J., McGlasson, W.B., Eaks, I.L., 1972. Treatment of fruit with propylene gives information about the biogenesis of ethylene.
Nature, 237(5352):235-236.
[22] Moon, H., Callahan, A.M., 2004. Developmental regulation of peach ACC oxidase promoter-GUS fusions in transgenic tomato fruits.
J Exp Bot, 55(402):1519-1528.
[23] Oeller, P.W., Lu, M.W., Taylor, L.P., 1991. Reversible inhibition of fruit senescence by antisense RNA.
Science, 254(5030):437-439.
[24] Ozga, J.A., Reinecke, D.M., 2003. Hormonal interactions in fruit development.
J Plant Growth Regul, 22(1):73-81.
[25] Pan, G., Lou, C., 2008. Isolation of an 1-aminocyclopropane-1-carboxylate oxidase gene from mulberry (
Morus alba L.) and analysis of the function of this gene in plant development and stresses response.
J Plant Physiol, 165(11):1204-1213.
[26] Ren, J., Leng, P., 2010. Role of abscisic acid and ethylene in fruit maturation of sweet cherry.
Acta Hort Sin, (in Chinese),37(2):199-206.
[27] Rieu, I., Cristescu, S.M., Harren, F.J.M., 2005.
Rp-ACS1, a flood-induced 1-aminocyclopropane-1-carboxylate synthase gene of
Rumex palustris, is involved in rhythmic ethylene production.
J Exp Bot, 56(413):841-849.
[28] Sebasti, C.H., Hardin, S.C., Clouse, S.D., 2004. Identification of a new motif for CDPK phosphorylation
in vitro that suggests ACC synthase may be a CDPK substrate.
Arch Biochem Biophys, 428(1):81-91.
[29] Seymour, G.B., Taylor, J.E., Tucke, G.A., 1993.
Biochemistry of Fruit Ripening, Chapman & Hall, Springer Netherlands,:
[30] Sun, L., Zhang, M., Ren, J., 2010. Reciprocity between abscisic acid and ethylene at the onset of berry ripening and after harvest.
BMC Plant Biol, 10(1):257
[31] Sunako, T., Sakuraba, W., Senda, M., 1999. An allele of the ripening-specific 1-aminocyclopropane-1-carboxylate synthase gene (
ACS1) in apple fruit with a long storage life.
Plant Physiol, 119(4):1297-1303.
[32] Tatsuki, M., Mori, H., 2001. Phosphorylation of tomato 1-amino-cyclopropane-1-carboxylic acid synthase, LE-ACS2, at the C-terminal region.
J Biol Chem, 276(30):28051-28057.
[33] Tatsuki, M., Haji, T., Yamaguchi, M., 2006. The involvement of 1-aminocyclopropane-1-carboxylic acid synthase isogene,
Pp-ACS1, in peach fruit softening.
J Exp Bot, 57(6):1281-1289.
[34] Tsuchisaka, A., Theologis, A., 2004. Heterodimeric interactions among the 1-amino-cyclopropane-1-carboxylate synthase polypeptides encoded by the
Arabidopsis gene family.
PNAS, 101(8):2275-2280.
[35] Varanasi, V., Shin, S., Mattheis, J., 2011. Expression profiles of the
MdACS3 gene suggest a function as an accelerator of apple (
Malus×
domestica) fruit ripening.
Posth Biol Technol, 62(2):141-148.
[36] Wang, A., Yamakake, J., Kudo, H., 2009. Null mutation of the
MdACS3 gene, coding for a ripening-specific 1-aminocyclopropane-1-carboxylate synthase, leads to long shelf life in apple fruit.
Plant Physiol, 151(1):391-399.
[37] Wang, K.L.C., Li, H., Ecker, J.R., 2002. Ethylene biosynthesis and signaling networks.
Plant Cell, 14(Suppl.):S131-S151.
[38] Xie, Y.H., Zhu, B.Z., Yang, X.L., 2006. Delay of postharvest ripening and senescence of tomato fruit through virus-induced
LeACS2 gene silencing.
Posth Biol Technol, 42(1):8-15.
[39] Yang, S.F., Hoffmann, N.E., 1984. Ethylene biosynthesis and its regulation in higher plants.
Annu Rev Plant Physiol, 35(1):155-189.
[40] Zaharah, S.S., Singh, Z., Symons, G.M., 2013. Mode of action of abscisic acid in triggering ethylene biosynthesis and softening during ripening in mango fruit.
Posth Biol Technol, 75:37-44.
[41] Zhang, M., Yuan, B., Leng, P., 2009. The role of ABA in triggering ethylene biosynthesis and ripening of tomato fruit.
J Exp Bot, 60(6):1579-1588.
[42] Zhang, Z., Ren, J.S., Clifton, I.J., 2004. Crystal structure and mechanistic implications of 1-aminocyclopropane-1-carboxylic acid oxidase—the ethylene-forming enzyme.
Chem Biol, 11(10):1383-1394.




Open peer comments: Debate/Discuss/Question/Opinion
<1>