1. Compositions and functions of polycomb group (PcG) complexes
PcG proteins were originally identified as a set of genes controlling proper body segmentation in
Drosophila via repressing
Hox genes (Lewis,
1978; Struhl,
1981). PcG proteins are now widely recognized in all metazoans for their roles in the variety of biological processes, such as cell cycle control, X-inactivation, maintenance of pluripotency and self-renewal in embryonic stem cells (ESCs), cell fate decisions, and developmental controls (Fig.
1c) (Muller and Verrijzer,
2009; Richly et al.,
2011; Aloia et al.,
2013). PcG proteins are assembled into a variety of multi-protein complexes, and the best characterized are polycomb repressive complexes 1 and 2 (PRC1 and PRC2). The PRC2 core complex of
Drosophila includes the enhancer of zeste [E(z)], the suppressor of zeste [Su(z)], and extra sex combs (Esc). The mammalian PRC2 core components comprise EZH1 or EZH2 [a homolog of E(z)], SUZ12 [a homolog of Su(z)], and an Esc homolog called EED. As the only PRC2 subunit with enzymatic activity, EZH2 is responsible for catalyzing the di- and tri-methylation of Lys27 on histone H3 (H3K27me
2 or 3) (Margueron et al.,
2008). The other core PRC2 components are necessary for complex assembly and for proper enzymatic activity (Pasini et al.,
2004; Margueron et al.,
2009). In
Drosophila, PRC1 core complex consists of polycomb (Pc), polyhomeotic (Ph), posterior sex combs (Psc), and sex combs extra (Sce) (also known as RING) (Fig.
1a), and mammals have several homologs to each subunit (Fig.
1b). There are five Pc (CBX2, CBX4, CBX6, CBX7, and CBX8), two Sce (RING1/RING1A and RING2/RING1B), three Ph (PHC1, PHC2, and PHC3), and six Psc known collectively as polycomb group ring fingers (PCGFs) (Morey and Helin,
2010).
Fig.1
Compositional diversity and functions of PRC1
(a) Main compositions of PRC1 in fruit flies; (b) Corresponding homologs of fly PRC1 subunits in humans; (c) Main functions known about PRC1. * The assembly of RYBP/YAF2 and CBX proteins in PRC1 may be mutually exclusive
For PRC1, recent advances indicate that the composition and patterns of assembly are far more biochemically diverse than originally estimated. Besides the canonical subunits listed above, there are many other PRC1 components and supporting factors, such as RING1/YY1-binding protein (RYBP) and its homolog YAF2, and the mammalian orthologs of the
Drosophila sex comb on midleg (Scm). For details about the complexity and diversity of the PRC1 subcategories, please refer to some recent reports and reviews (Gao et al.,
2012; Schwartz and Pirrotta,
2013). The compositions among different mammalian PRC1 subcategories are largely divergent while all of these PRC1 complexes contain at least one of the two RING proteins, which are known as an E3 ubiquitin ligase responsible for ubiquitination of histone H2A (H2AK119ub) (Wang et al.,
2004). Several types of PRC1 have been shown to have none or low levels of chromobox (CBX) proteins. RYBP or YAF2 is known to stimulate the enzymatic activity of RING1B toward H2AK119ub, while CBX proteins may not exist simultaneously in this type of PRC1 (Gao et al.,
2012).
2. CBX proteins in PRC1
There are at least eight members of the CBX proteins in both mouse and human genomes, each of which contains a single N-terminal chromodomain (Wotton and Merrill,
2007). They are all involved in the regulation of heterochromatin, gene expression, and developmental programs. They are further divided into two groups: (1) CBX1, CBX3, and CBX5, also known as heterochromatin protein 1β (HP1β), HP1γ, and HP1α, respectively; (2) CBX2, CBX4, CBX6, CBX7, and CBX8, all having a C-terminal polycomb repressor box, serve as canonical components in PRC1 (Wotton and Merrill,
2007). In this review, we will mainly discuss the roles of these five CBX proteins in the context of PRC1.
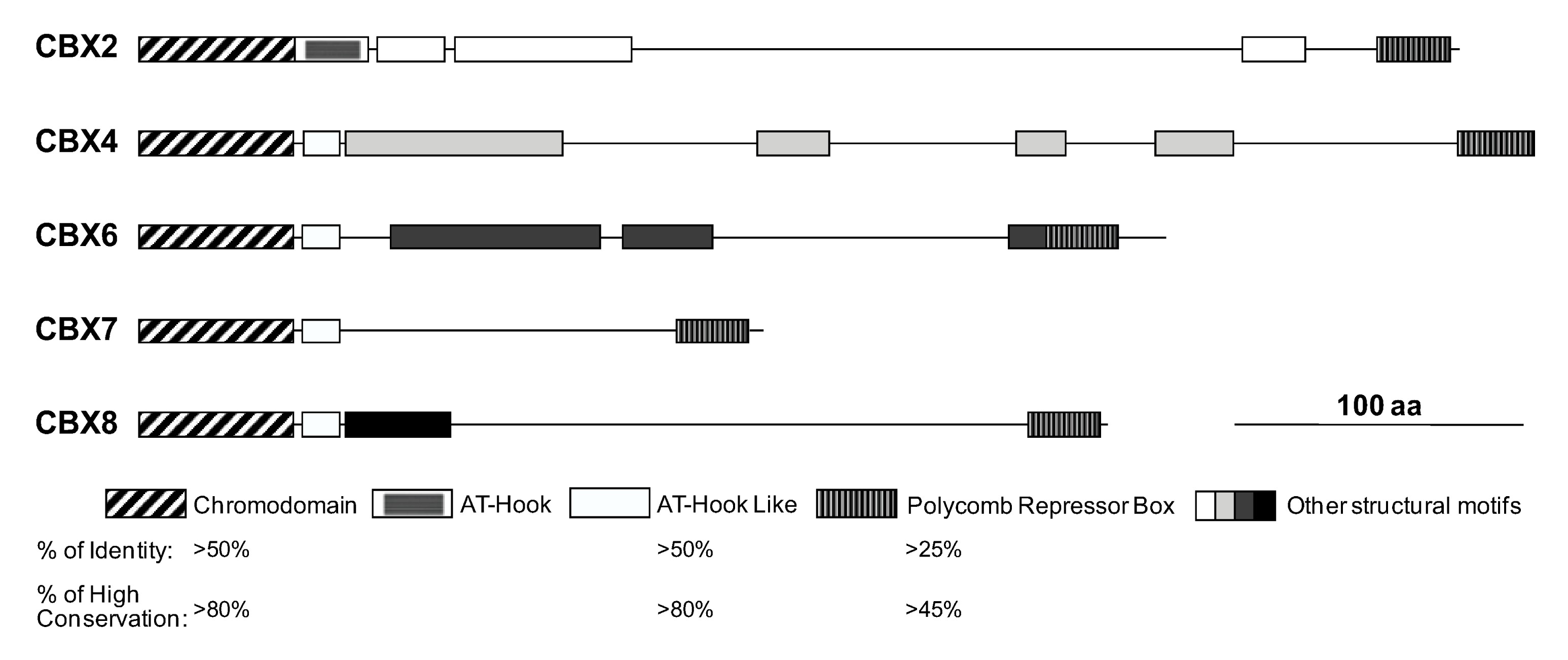
The domain organization of CBX proteins is shown in Fig.
2. CBX proteins physically interact with H3K27me
3 via their chromodomains, which helps the recruitment and stabilization of PRC1 to specific regions of the chromatin (Bernstein B.E. et al.,
2006; Buchwald et al.,
2006). It was reported that mammalian CBX proteins exhibited differential bindings to methylated histone tails. CBX2 and CBX7 bind to both H3K9Me
3 and H3K27Me
3 whereas CBX4 shows stronger affinity for H3K9Me
3 (Bernstein E. et al.,
2006). The C-terminal polycomb repressor box of CBX proteins is involved in transcriptional silencing and binding to other PRC1 components such as RING1B (Muller et al.,
1995; Bezsonova et al.,
2009). Adjacent to chromodomain, all these CBX homologs have a DNA binding motif, AT-hook (in CBX2) or an AT-hook like motif (in the other four CBX proteins) (Senthilkumar and Mishra,
2009). The less conserved sequences in the middle of the CBX proteins may play a role in specifically directing each CBX family member to distinct regions of the chromatin (Vincenz and Kerppola,
2008).
Fig.2Domain organization and conservation among CBX proteins
% of identity: ratio of the number of identical amino acid (aa) to the number of total aa; % of high conservation: ratio of the number of highly conserved aa to the number of total aa. This figure is modified from Senthilkumar and Mishra (
2009)
3. Recruitment of PcG complexes
PcG complexes are involved in repressing over several thousand genes in mammalian genomes and the pool of their target genes is distinct in different cell types and dynamically changing in various cell states (Simon and Kingston,
2013). Components in PRC1 and PRC2 generally do not have DNA binding properties. The targeting mechanisms of the mammalian PcG complexes are diverse and extremely complicated. Many sources of the ‘targeting command’ are comprehensively integrated, including chromatin signatures (such as histone modifications, histone variants, DNA sequences, and CpG islands), varieties of non-coding RNA (ncRNA) species, transcription factors and cofactors, and possibly the status of RNA polymerase II (Pol II).
In
Drosophila, specific DNA modules called polycomb response elements (PREs) are responsible for recruiting PcG complexes to their chromatin targeting regions (Simon et al.,
1993). This process is mediated by sequence-specific DNA binding proteins such as the zinc finger protein PHO. Recent studies found that
Drosophila PREs consist of binding sites for a complex array of DNA binding factors and the interplay among these regulatory factors is important for specifying the function of the PREs in a cell or tissue-specific fashion (Oktaba et al.,
2008; Brown and Kassis,
2013). In mammals, it becomes much more difficult in searching and defining ‘PRE’. Transcription repressor protein YY1 is the mammalian homolog of the
Drosophila PHO, which works together with YAF2 and the transcription corepressor C-terminal binding protein (CtBP) in recruiting PcG complexes to many of their target genes, including
HOX genes (Atchison et al.,
2003; Srinivasan and Atchison,
2004; Basu et al.,
2014). Several potential human PREs have been found in
HOXB,
HOXC, and
HOXD clusters to facilitate the recruitment of both PRC2 and PRC1 components to a reporter gene, yet the DNA binding factors mediating these interactions are variable (Woo et al.,
2010;
2013). Another potential human PRE containing 25 repeats of YY1 binding motif was validated to be able to down-regulate reporter genes via PRC2 in both HEK cells and in
Drosophila (Bengani et al.,
2013). Besides YY1, many other transcription factors have been reported to act as recruiters for PcG complexes, such as neuronal inhibitor REST (Ren and Kerppola,
2011).
In addition to potential PREs, chromatin regions enriched in CpG dinucleotides (CpG islands) may also facilitate targeting of PcG complexes in mammals. CpG islands that lack both 5-methylcytosine and activator-binding sites are largely overlapping with H3K27me
3, PRC2, and PRC1, and notably, constructs, containing CpG island-like sequences, are found to be capable of recruiting PRC2 upon integration into a mouse genome (Ku et al.,
2008; Mendenhall et al.,
2010). One PRC1 component Kdm2b/Fbxl10 was recently identified as a binding factor towards CpG islands (Wu X. et al.,
2013). Other signatures of the chromatin may also contribute to the recruitment of PcG complexes, such as histone variants H2AZ (Creyghton et al.,
2008) and MacroH2A (Buschbeck et al.,
2009).
The accumulating evidence has emphasized another scheme involved in ncRNA for the recruitment of PcG complexes. In the model of mammalian X chromosome inactivation, the long non-coding RNA (lncRNA) Xist (more specifically its A repeat region called RepA) has been implicated in PRC2 recruitment (Plath et al.,
2003; Zhao et al.,
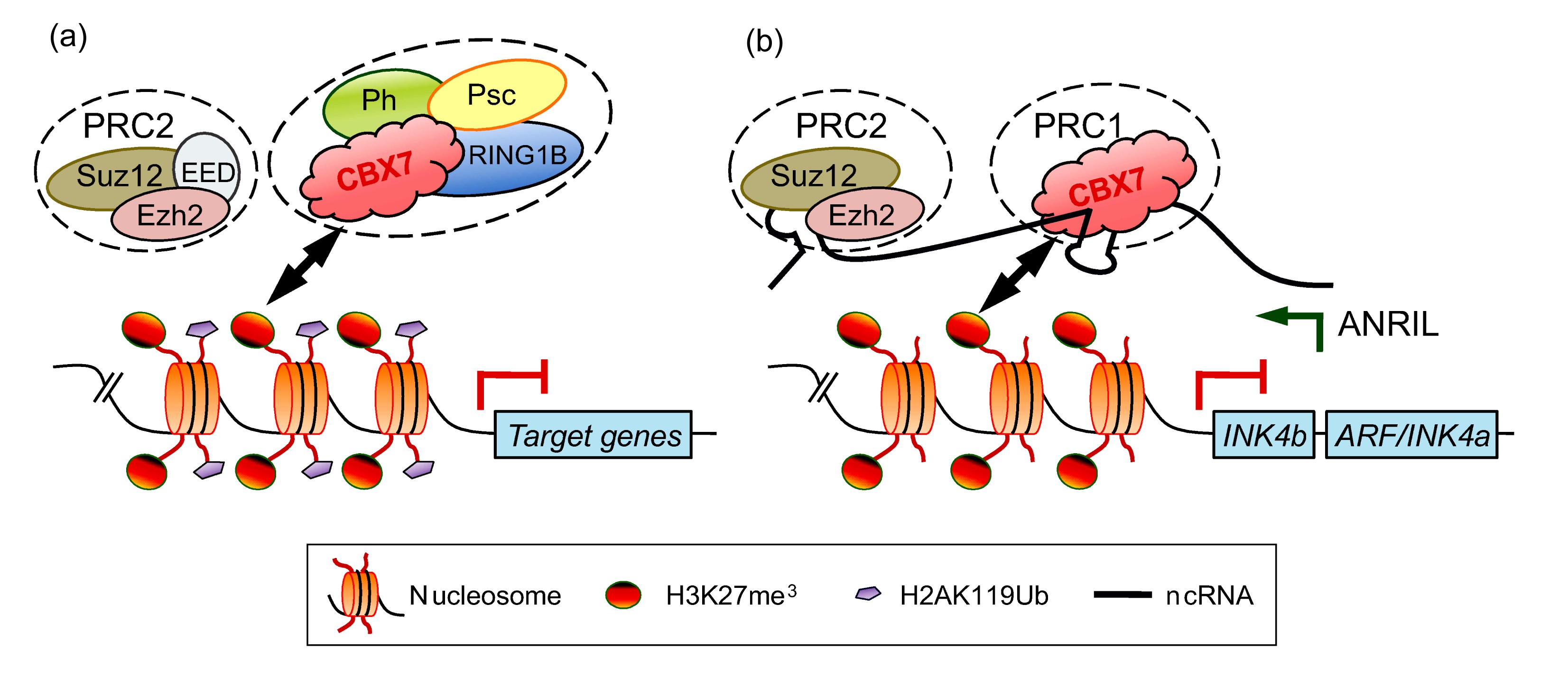
2008). Both Suz12 and Ezh2 are known to have RNA binding capability in PRC2. The chromatin recruitment of canonical PRC1 complex is mainly mediated by CBX proteins. As mentioned above, the histone modification mark H3K27me
3 serves as a signal for CBX protein mediated PRC1 recruitment (Fig.
3a). CBX proteins are the only known RNA binding proteins in PRC1 and their chromodomains are required for RNA binding (Bernstein E. et al.,
2006). At the
Ink4a/
ARF/
Ink4b locus, the anti-sence lncRNA ANRIL has been demonstrated to recruit both PRC1 and PRC2 in a
cis-manner, and this guidance plays a key role in repressing these target genes by PcG complexes (Yap et al.,
2010; Kotake et al.,
2011) (Fig.
3b). Other similar examples including H19 ncRNA and Kcnqtlot1 ncRNA are described in more detail in other reviews (Brockdorff,
2013; Simon and Kingston,
2013). ncRNA may also recruit PcG complexes in a
trans-manner. The only reported example is the human HOTAIR ncRNA, which is transcribed from the
HOXC loci while having influence on genes located in the
HOXD loci (Tsai et al.,
2010). However, deletion of the putative mouse HOTAIR did not exhibit any effects on PRC2 targeting (Schorderet and Duboule,
2011), and deleting various segments within the mouse
HoxD loci did not disturb the overall patterns of PRC2 recruitment (Schorderet et al.,
2013). Besides lncRNA species, genome-wide studies have revealed that promoter-associated short RNAs transcribed from CpG island promoters and other types of RNA species also potentially facilitate the recruitment of PRC2 (Kanhere et al.,
2010; Zhao et al.,
2010).
Fig.3
Recruitment of PRC1 to chromatin through CBX proteins
Recruitment through binding to histone marks H3K27me3 (a) and/or binding to ncRNA (b). Note that only RNA binding components in each complex are shown in (b)
In addition to the above mechanisms, the phosphorylation status of the serine residues within the carboxyl-terminal domain (CTD) of Pol II has also been shown to correlate with the occupancy of PcG complexes at certain groups of target genes. The phosphorylation at Ser5 normally occurs right after initiation (Ser5p
+) while the phosphorylation at Ser2 and Ser7 positions are generally associated with productive elongation. The new genome-wide study discovered that in mouse ESC, the development-related PRC targets were generally associated with poised Pol II (Ser5p
+, Ser2p
−, Ser7p
−) while PRC targets involved in metabolism were decorated with transcriptional active Pol II (Ser5p
+, Ser2p
+, Ser7p
+) (Brookes et al.,
2012). It remains highly debatable whether the chromatin-bound PcG complexes help establish the block of transcription elongation or the preset states of Pol II serve as signs for recruiting the PcG complex.
Although the targeting of PRC1 to chromatin is mainly dependent on PRC2, differential recruitment of mammalian PRC1 vs. PRC2 has been observed in several cases (Ren et al.,
2008; Vincenz and Kerppola,
2008; Tavares et al.,
2012; Wu X. et al.,
2013). The detailed mechanisms remain unclear. PRC1 may be initially recruited to chromatin regions distal to the target genes, searching for its targets; further recognition to H3K27me
3 with or without the guidance of ncRNA helps direct and stabilize the complex to final destinations.
4. Molecular functions of CBX proteins
4.1. Functions in common
The functions of CBX family proteins have been studied in many cell types. The
Ink4a/
ARF/
Ink4b locus is one of the earliest identified and the most well-known targets for CBX-containing PRC1 complex (Gil et al.,
2004). The three tumor suppressors encoded by this locus play a central role in cell-cycle inhibition, senescence, and stress-induced apoptosis. Consistently, knockdown of
Cbx genes is generally associated with reduced cell proliferation, and in contrast, overexpression of
Cbx genes in some primary cells normally results in the extension of life span or even cell immortalization (Gil et al.,
2004) (Table
1).
Table 1
Effects of knockdown or overexpression of Cbx genes
| Gene |
Cell type |
Species |
Phenotype |
Reference |
| Knockdown |
| Cbx2
|
HSC and progenitor cell |
Human |
Decreased proliferation and increased apoptosis; increased expression of the cyclin-dependent kinase inhibitor p21 |
van den Boom et al. (2013) |
| HSC |
Mouse |
No effect on HSC self renewal |
| Cbx4
|
Epidermal stem cell |
Human |
Reduced size and number of macroscopic colonies; terminal differentiation; increased size and granularity of differentiated cells |
Luis et al. (2011) |
| HCC |
Human |
Decreased cell proliferation and slower cell cycle progression |
Wang et al. (2013) |
| Cbx7
|
Primary and cancerous prostate cell lines |
Human |
Impairment of cell growth via controlling Ink4a/Arf genes |
Bernard et al. (2005) |
| ESC |
Mouse |
Differentiation; de-repression of PRC1 target genes, particularly ectoderm lineage associated genes |
Morey et al. (2012); O′Loghlen et al. (2012) |
| Cbx8
|
Intervertebral disc nucleus pulposus cell |
Rat |
Decreased proliferation |
Zhou et al. (2013) |
|
| Overexpression |
| Cbx2
|
Hematopoietic stem/progenitor cell |
Mouse |
Differentiation |
Klauke et al. (2013) |
| Cbx4
|
Epidermal stem cell |
Human |
Colonies remain undifferentiated but smaller |
Luis et al. (2011) |
| HSC |
Mouse |
Differentiation; decreased proliferation; lower numbers of secondary colonies |
Klauke et al. (2013) |
| Cbx6
|
GBM cell |
Human |
Inhibited proliferation |
Li et al. (2013) |
| Cbx7
|
Keratinocytes (NHKs), fibroblast cell line |
Human |
Extended replicative capacity (no immortalization) |
Gil et al. (2004) |
| MEF |
Mouse |
Extended replicative capacity; immortalization |
Gil et al. (2004) |
| Prostate cancer cell line (e.g., LNCaP) |
Human |
Slight growth advantage in both androgen-dependent and independent conditions |
Bernard et al. (2005) |
| HSC or early progenitor cells |
Mouse |
Produced transgenic chimeric mice developed tumors within one year of adoptive transfer; enlarged thymus and splenomegaly; promoted T cell lymphomas genesis; cooperated with c-myc to produce highly aggressive B-cell lymphomas |
Scott et al. (2007) |
| Thyroid carcinoma cell lines (e.g., ARO) |
Human |
Lower growth potential; less colony formation |
Pallante et al. (2008) |
| EC (TERA-2) |
Human |
Growth advantage; resistance to retinoic acid-induced differentiation |
Mohammad et al. (2009) |
| CRC cell lines |
Human |
Decreased proliferation |
Pallante et al. (2010) |
| ESC |
Mouse |
Lower level of spontaneous differentiation |
O′Loghlen et al. (2012) |
| Cbx8
|
HSC |
Mouse |
Differentiation; decreased proliferation; less secondary colonies |
Klauke et al. (2013) |
CRC: colorectal cancer; EC: embryonal carcinoma cell; ESC: embryonic stem cell; GBM: glioblastoma multiforme; HCC: hepatocellular carcinoma cell line; HSC: hematopoietic stem cell; MEF: mouse embryonic fibroblast; NHKs: normal human keratinocytes
4.2. Interplay among CBX proteins
In stem cells, the consequences upon changing expressions of various CBX family members are not very consistent because it seems that CBX/PRC1 can promote both self-renewal and differentiation. This apparent inconsistency can now be better explained by recent discoveries of an autoregulatory loop among CBX family members (Morey et al.,
2012; O′Loghlen et al.,
2012; Klauke et al.,
2013) (Fig.
4a). Studies indicated that CBX7 was the primary CBX protein assembled in PRC1 in the mouse ESC, and notably, it directly participated in transcription repression of the genes encoding other CBX family members (including
Cbx2,
Cbx4, and
Cbx8) by PcG complexes. Interestingly, there was a dynamic switch of the identity of CBX proteins in PRC1 upon differentiation. During ESC differentiation, the expression of
Cbx7 was down-regulated and
Cbx2,
Cbx4, and
Cbx8 were up-regulated simultaneously. The mechanism that triggers the onset of this switch remains elusive. Depletion of CBX7 induced differentiation of ESC and mainly ectoderm-lineage-associated genes were derepressed, while the ectopic expression of
Cbx7 enhanced ESC self-renewal. In differentiating cells, CBX2 and CBX4 replaced the function of CBX7 in PRC1 targeting but they were found to have non-overlapping functions, repressing distinct target genes. The teratomas derived from CBX2- and CBX4-depleted ESC displayed abnormal differentiation phenotypes with increased number of cells expressing endodermal or mesodermal marker genes. More recent studies demonstrated that in mouse hematopoietic stem cells (HSCs), CBX7 was also required for their self-renewal, whereas CBX2, CBX4, and CBX8 were crucial for their proper differentiation (Klauke et al.,
2013). Ectopically expressed CBX2, CBX4, or CBX8 in HSC was able to compete with CBX7 in PRC1 assembly while they shifted the targets of PRC1 and resulted in the differentiation of HSC. However, in a human study, the depletion of CBX2 resulted in decreased cell proliferation and increasing levels of apoptosis, emphasizing the requirement of CBX2 in the maintenance of HSC (van den Boom et al.,
2013). Overall, in stem cells (including embryonic and adult stem cells), CBX proteins play a key role in maintenance of pluripotency or multipotency. The discrepant results in some of these studies suggest the specific roles each CBX protein might be variable in different species. Some data about manipulation of
Cbx expression in cancer cell lines are also listed in Table
1, while the roles of CBX proteins in cancer will be mainly discussed later in this review.
Fig.4
Interplay among CBX proteins
(a) Auto-regulation among CBX proteins in proliferating vs. differentiated mouse ESCs (mESCs); (b) Co-function among CBX proteins in human fibroblasts
Besides the mutually inhibitory mode described above, multiple CBX proteins can also be expressed and shown to be functioning in the same cell type. Through genome-wide analyses, Klauke et al. (
2013) found that targets of CBX7 and CBX8 were largely overlapping in the mouse HSC. Meanwhile, they also identified about 200 genes that were differentially targeted. Interestingly, the genes uniquely occupied by CBX8 were actually highly expressed in HSC and later became repressed in progenitors, which was opposite to the expression pattern of the CBX7 targets. This suggests that CBX/PRC1 can be pre-deposited to the future targets and distinct CBX family members may help specify the ‘actively repressed’ vs. ‘ready-to-be repressed’ targets. A similar phenomenon was also observed by Pemberton et al. (
2014). In this study, the genome-wide occupancy of several PRC1 components was analyzed and compared in human fibroblast cell lines. The results demonstrated that the target genes of CBX6, CBX7, and CBX8 were mostly overlapping, yet the detailed configuration of co-occupied PRC1 complexes remains unknown (Pemberton et al.,
2014). Theoretically, PRC1 complexes containing each CBX protein respectively could be lined up or form oligomers at their common targets, or it is possible that various CBX proteins are integrated into a single unit of PRC1 complex (Fig.
4b). Notably, Pemberton et al. (
2014) also identified that cell type-specific PRC1 binding sites and the occupancy of PRC1 at many loci did not correlate with an outcome of transcription repression.
4.3. Phenotypes of Cbx knockout (KO) mice
The roles of CBX proteins playing during embryogenesis have been investigated in KO mice or human patients with genetic mutations (Table
2). The observed phenotypes revealed the functional specification of these CBX family members. All known
Cbx KO mice (
Cbx2,
Cbx4, and
Cbx7) can survive the entire embryonic development. Homozygous mutants of
Cbx2 or
Cbx4 KO mice displayed postnatal lethality. The distinct features of
Cbx2 KO mice include male-to-female sex reversal and defects related to skeletal development were observed in two separate KO lines. Some of these functions of CBX2 are apparently conserved across species: a combination of two point mutations in
Cbx2 led to male-to-female sex reversal in humans without other obvious developmental defects. CBX2 plays a critical role in germ cell development, meiosis onset and homologous chromosome synapsis in the mammalian germ line (Baumann and de la Fuente,
2011). CBX4 has been recently found to play a key role in the development of the immune system: the proliferation of thymic epithelial cells and the maintenance of thymic epithelium were impaired in the KO mice. The
Cbx7 KO mice were grossly normal in morphology and growth although these mice attempted to develop liver and lung adenomas and carcinomas at adulthood. Considering the dominant role it plays in mouse ESC compared to other CBX proteins, alternative pathways have to be applied to compensate for the loss of CBX7 in these mice.
Table 2
Phenotypes of Cbx knockout mice and human patient with point mutations
| Gene |
Species |
Phenotypes of gene knockout mice or human patient with point mutations |
Note |
Reference |
|
Cbx2
|
Mouse |
50% −/− mutants died within a few hours after born; 90% mutants died within 4 weeks and the lifespan did not exceed 6 weeks; a malformation of the exoccipital bone and the deficiency of a cervical vertebrae and a vertebrosternal rib; aberrant T cell expansion; senescence of embryonic fibroblasts |
Excision of exons 1–4 (aa 1–96) |
Coré et al. (1997; 2004) |
| Mouse |
About 30% of XY−/− embryos displayed sex reversal; recovered germ cells exhibited premature meiosis onset; XX−/− had small ovaries with severe germ cell loss; abnormal chromosome synapsis and structural damage of chromosomes during meiosis |
Excision of exons 1–4 (aa 1–96) |
Baumann and de la Fuente (2011) |
| Mouse |
50% −/− mutants died within 2 weeks after born; male survivors showed male-to-female sex reversal, and female survivors exhibited two smaller ovaries or the absence of an ovary; transformation of the axial skeleton and hypoplastic gonad formation; adrenal and spleen hypoplasia; male-to-female sex reversal rescued by crossing them with transgenic mice displaying forced expression of Sry or Sox9; yet testes remained hypoplastic |
Disruption within exon 5 (affected aa 159–519) |
Katoh-Fukui et al. (1998; 2005; 2012) |
| Human |
Male-to-female sex reversal: a girl (with uterus and histologically normal ovaries) displays 46, XY karyotype; the girl had normal female internal and external genitalia and normal bilateral ovaries instead of dysgenetic gonads at histology; no obvious defects related to skeletal development, adrenal, or splenic growth |
Two point mutations (P98L and R443P) |
Biason-Lauber et al. (2009) |
|
Cbx4
|
Mouse |
+/− mice: grossly normal in morphology and growth; −/− mutant mice: born alive at an expected Mendelian ratio but died within 1 h of birth; severely retarded growth of the mutant thymus; decreased cell numbers of both total thymic cells and thymic epithelial cells; numbers of thymocytes (E17.5) increased in G0/G1 phase, and reduced in S and G2/M phases |
Excision of exons 1–2 (aa 1–38) |
Liu et al. (2013) |
|
Cbx7
|
Mouse |
Increased body length for both sexes; mutants tend to develop liver and lung adenomas and carcinomas at adulthood; MEF cells from KO mice grow faster than WT controls |
Excision of exons 5–6 (aa 83–158) |
Forzati et al. (2012a) |
aa: amino acid; MEF: mouse embryo fibroblast; WT: wild type; −/−: homozygous knockout; +/−: heterozygous knockout
4.4. Unique roles of CBX proteins
In the five CBX proteins, CBX4 is the only one known to have an enzymatic activity. It acts as a small ubiquitin-like modifier (SUMO) E3 ligase and this activity is dependent on its chromodomain and a C-terminal substrate binding domain (Kagey et al.,
2003). SUMOylation has been implicated in the regulation of many cellular processes, including transcriptional repression, genome stability, chromatin organization, and DNA repair (Galanty et al.,
2009; Morris et al.,
2009; Dou et al.,
2011). It has been well known that SUMO is covalently conjugated to lysine residues on its substrates while there are also specific motifs identified in SUMO E3 ligases that mediate non-covalent interactions with SUMO (Song et al.,
2004). CBX4 indeed contains two such motifs that facilitate non-covalent SUMO binding and are required for full E3 ligase activity (Nacerddine et al.,
2005). The known substrates for CBX4 are listed in Table
3. The CBX4-mediated SUMOylations of CCCTC-binding factor (CTCF) and homeo-domain interacting protein kinase 2 (HIPK2) are associated with transcriptional repression. The SUMOylations of heterogeneous nuclear ribonucleoprotein K (HnRNP-K) and BMI1 are essential for their stabilization or site-specific targeting, respectively, in response to DNA damage.
Table 3
SUMOlyation substrates of CBX4
| Substrate |
Site of Rxn |
Molecular function |
Reference |
| CtBP |
K428 |
Make the SUMOylated CtBP preferentially associate with CBX4 |
Kagey et al. (2003) |
| SIP1 |
K391 and K866 |
Attenuate its transcriptional repression activity |
Long et al. (2005) |
| HIPK2 |
K25 |
Enhance the ability of HIPK2 to mediate transcriptional repression |
Roscic et al. (2006) |
| Dnmt3a |
An extended PWWP region |
Involved in the functional regulation of DNA methyltransferases by promoting their SUMO modification |
Li et al. (2007) |
| CBS |
Not known |
Decrease CBS activity of catalyzing the first irreversible step in the transsulfuration pathway |
Agrawal and Banerjee (2008) |
| CTCF |
K74 and K698 |
Contribute to the repressive function of CTCF on the c-myc P2 promoter |
MacPherson et al. (2009) |
| E2F1 |
K266 |
Required for the activation of cell-growth-control genes in response to serum |
Yang et al. (2011) |
| α-Synuclein |
Not known |
Function as a cytoprotector by increasing α-synuclein aggregate formation within fibroblast cells |
Oh et al. (2011) |
| HnRNP K |
K422 |
Required for p53-mediated cell-cycle arrest in upon DNA damage |
Lee et al. (2012); Pelisch et al. (2012) |
| ZNF131 |
K567 |
Potentiate the inhibitory effect of ZNF131 on estrogen signaling and attenuate estrogen-induced cell growth in MCF-7 cells |
Oh and Chung (2012) |
| BMI1 |
K88 |
Essential for BMI1 targeting to the sites of DNA breaks |
Ismail et al. (2012) |
CBS: cystathionine-β-synthase; CTCF: CCCTC-binding factor; Dnmt3a: DNA methyltransferase 3a; HIPK2: homeodomain interacting protein kinase 2; HnRNP-K: heterogeneous nuclear ribonucleoprotein-K; SIP1: Smad-interacting protein 1; ZNF131: zinc finger protein 131
Like other CBX proteins, CBX8 was found in diploid human and mouse fibroblasts to regulate premature senescence through controlling the transcription of the
Ink4a/
Arf locus (Dietrich et al.,
2007). Recently, Zhou et al. (
2013) discovered that the DN oxidative damage caused up-regulation of
Cbx8 expression and blocking this response by silencing
Cbx8 led to more serious DNA damage, indicating that CBX8 played an important role in DNA repair. Notably, some recent studies uncovered a novel inhibitory mechanism for PRC1 activity. CBX8 acts as an essential cofactor required for mixed lineage leukemia (MLL)-AF9 fusion or MLL-ENL fusion-induced transcriptional activation and leukemic transformation, which is contrary to its role as a transcriptional repressor in PRC1 (Tan et al.,
2011; Maethner et al.,
2013). The physical interaction between ENL and CBX8 eliminated the transcription repression activity of PRC1, which was required for efficient transformation of hematopoietic cells (Maethner et al.,
2013). Along with that, CBX8 competed with H3K79 methyltransferase disruptor of telomeric silencing 1-like (DOT1L) for binding to AF9, resulting in up-regulation of the
ENaCα gene, a known target of AF9-DOT1L (Malik and Hemenway,
2013). In this case, it was known that CBX8 worked in the context of PRC1 since the other PRC1 components including RING1B and BIM1 were found together with CBX8. Although essential for MLL-AF9 induced leukemic transformation, analyses of the
Cbx8 conditional-depletion mice on hematopoietic steady-state conditions revealed that CBX8 is not required for steady-state hematopoiesis, long-term HSC maintenance, or stem and progenitor cell function (Tan et al.,
2011).
5. Regulations of CBX proteins
Besides the auto-regulation among CBX family members at the transcriptional level discussed above, CBX proteins are also subjected to various post-translational regulations. The reported information about identified post-translational modifications is listed in Table
4, which involves phosphorylation, SUMOylation/de-SUMOylation, and methylation/de-methylation. Although phosphorylation events have been detected in all five CBX proteins, it is not clear whether they share any common functional mechanisms. Many phosphorylation sites were detected in mass spectrometry analyses without further validation. The most recent discovery was that phosphorylation of CBX7 by mitogen-activated protein kinase (MAPK) helped strengthen its association with PRC1 upon mitogen stimulation. As a SUMO E3 ligase, CBX4 itself is subject to the reversible regulation of SUMOylation and de-SUMOylation, and SUMOylation at Lys492 facilitates its interaction with H3K27me
3 (Kang et al.,
2010). A recent work has demonstrated that the unmethylated and methylated forms of CBX4 specifically bind to distinct ncRNA named MALAT1/NEAT2 and TUG1, respectively (Yang et al.,
2011). The switch between these interactions correlates with the subnuclear relocation of growth-control genes in response to growth signals. The binding of NEAT2 to unmethylated CBX4 promotes E2F1 SUMOylation, resulting in activation of the growth-control genes. Some key points remain unclear, such as whether the enzymatic activity of CBX4 in this transcription activation event relies on the presence of PRC1 complex, and whether the outcome of transcription activation is achieved by somehow inhibiting the PRC1 function.
Table 4
Post-translational modification of CBX proteins
| Protein |
Modification |
Modifying enzyme |
Site |
Function of modification |
Reference |
| CBX2 |
Phosphorylation |
Casein kinase 2 (CK2) |
S42 |
Reduction of binding to H3K9me3 and increase of binding to H3K27me3 (in vitro) |
Hatano et al. (2010) |
| CBX4 |
SUMOylation |
CBX4 or unknown |
K492 |
Stimulation of its interaction to H3K27me3
|
Kang et al. (2010) |
| De-SUMOylation |
SENP2 |
K492 |
Reduction of its binding to H3K27me3
|
Kang et al. (2010) |
| Phosphorylation |
Akt1 |
S415, S434 |
Help integrate multiple enzymatic activities to target CtBP1 for degradation |
Merrill et al. (2010) |
| HIPK2 |
T495 |
Promote CBX4’s ability to increase HIPK2 SUMOylation |
Roscic et al. (2006) |
| Unknown |
Y205 |
Identified in proteomics study using lung cancer samples; mechanisms unknown |
Rikova et al. (2007) |
| Unknown |
S347, T349 |
Identified in proteomics study of cell cycle related changes; mechanisms unknown |
Dephoure et al. (2008) |
| Unknown |
S413 |
Identified in proteomics study of EGF-stimulated cell signaling networks; mechanisms unknown |
Olsen et al. (2006) |
| Unknown |
T415, T435 |
Identified in proteomics study searching for cellular targets of rapamycin in cancer cells; mechanisms unknown |
Chen et al. (2009) |
| Unknown |
S430 |
Identified in proteomics study of EGF-pathway; mechanisms unknown |
Cantin et al. (2008) |
| Methylation |
Suv39h1 |
K191 |
Methylated CBX4 acts as an important anti-mitogenic signal |
Yang et al. (2011) |
| De-methylation |
KDM4C |
K191 |
Unmethylated CBX4 is essential for physiological growth-control gene expression and cell proliferation |
Yang et al. (2011) |
| CBX6 |
Phosphorylation |
Unknown |
S107 |
Identified in proteomics study searching for cellular targets of rapamycin in cancer cells; mechanisms unknown |
Chen et al. (2009) |
| Unknown |
S301, S303 |
Identified in proteomics study of EGF-pathway; mechanisms unknown |
Cantin et al. (2008) |
| CBX7 |
Phosphorylation |
MAPK |
T118 |
Enhance its association with PRC1 upon mitogen stimulation. The specific molecular mechanisms unknown |
Wu H.A. et al. (2013) |
| CBX8 |
Phosphorylation |
Unknown |
S110, S130, S265, S352, S354, T234 |
Identified in proteomics study of cell cycle related changes; mechanisms unknown |
Daub et al. (2008); Dephoure et al. (2008); Chen et al. (2009) |
| Unknown |
S256 |
Identified in proteomics study searching for cellular targets of rapamycin in cancer cells; mechanisms unknown |
Daub et al. (2008); Chen et al. (2009) |
| Unknown |
S332 |
Identified in proteomics study of EGF-pathway; mechanisms unknown |
Cantin et al. (2008); Dephoure et al. (2008) |
EGF: epidermal growth factor; SENP2: SUMO-specific protease 2; MAPK: mitogen-activated protein kinase
6. Misregulation of CBX proteins and cancer
Misregulation of PcG proteins has been associated with many cancer types. In regard to CBX proteins, there is increasing evidence that they play an important role in tumor initiation, progression, and development by blocking differentiation and promoting self-renewal of cancer stem cells. The reports about misregulation of CBX proteins in various cancers are listed in Table
5. The findings are categorized based on the direction of expression changes for these CBX proteins in a given cancer type. CBX7 is the most characterized CBX protein in cancer-associated studies. The level of CBX7 is elevated in prostate, lymphoma, and gastric cancer; conversely loss of CBX7 has been detected in a prevalence of other cancer types, such as thyroid, colon, pancreatic, and breast cancer. The apparent dual activities of CBX proteins as either oncogenes or tumor suppressors in distinct cancer types may mirror their specialized function during embryonic development and tissue specification. As the discovery about mouse HSC discussed above, CBX7 was the primary CBX protein in the maintenance of the stemness and elevated level of CBX7 promoted T cell lymphomas genesis. However, in the same system, over-expression of other ‘non-dominant’ CBX proteins including CBX2, CBX4, and CBX8 actually triggered cell differentiation and decreased cell proliferation (Klauke et al.,
2013).
Table 5
Cancers associated with abberant expression of CBX proteins
| Expression |
Type of cancer |
Cancer cell line or specific sample |
Reference |
Molecular characterization |
| Elevated |
|
Cbx4
|
Hepatocellular carcinoma |
HCC cell lines (e.g., BEL-7042) |
Wang et al. (2013) |
Knockdown of Cbx4 resulted in decrease of both proliferating cell nuclear antigen (PCNA), cyclin E2, and increase of p16
|
|
Cbx7
|
Prostate cancer |
Prostate cancer cell lines (e.g., DU-145) |
Bernard et al. (2005) |
Repressed expression from Ink4a/Arf locus |
| Preneoplastic PIN and PCa prostate cancer cells |
Yap et al. (2010) |
| Lymphoma cancer |
Germinal center-derived follicular lymphomas |
Scott et al. (2007) |
| Gastric cancer |
Gastric cancer cell lines (e.g., SGC-7901) |
Zhang et al. (2010) |
|
Cbx8
|
GBM |
GBM cell lines (e.g., T98G) |
Li et al. (2013) |
Slightly increased expression (<2 fold); mechanisms not clear |
| Breast cancer |
Breast cancer cell lines (e.g., MCF7) |
Lee et al. (2013) |
Cooperated with SIRT1 for suppressing p53 acetylation; repressed the expression of p21 by inhibiting p53 binding to the promoter |
|
| Declined |
|
Cbx6
|
GBM |
GBM cell lines (e.g., T98G) |
Li et al. (2013) |
Decreased proliferative capacity and growth arrest when over-expressed; mechanisms not clear |
|
Cbx7
|
Thyroid cancer |
Thyroid cancer cell lines (e.g., TPC-1) |
Pallante et al. (2008) |
Levels of CBX7 gradually decrease with the progression of cancer and correlate with the increase of Ink4a/Arf expression |
| Lung cancer |
MEF isolated fromCbx7 knockout mice |
Forzati et al. (2012a) |
Up-regulated expression of CCNE1 gene; competing with CCNE1 activator HMGA1 for binding |
| Colon cancer |
Colorectal cancer cell lines (e.g., HT29) |
Pallante et al. (2010) |
Loss of CBX7 expression correlates with a poor outcome of colorectal cancer; mechanisms not clear |
| Pancreatic cancer |
Samples from pancreatic cancer patients |
Karamitopoulou et al. (2010) |
Levels of CBX7 gradually decrease with the progression of cancer and correlate with the loss of E-cadherin |
| Urothelial carcinoma |
Samples from bladder cancer patients |
Hinz et al. (2008) |
Mechanisms not clear |
| Breast cancer |
Breast cancer cell lines (e.g., MCF7) |
Hannafon et al. (2011) |
Loss of the tumor suppressor miR-125b and gain of the oncogenic miRNAs (miR-182 and -183) |
| GBM |
GBM cell lines (e.g., T98G) |
Li et al. (2013) |
Mechanisms not clear |
| Ovarian clear cell adeno-carcinoma |
OCCA cell lines (e.g., TOV21G/KOC-7C) |
Shinjo et al. (2013) |
Up-regulated expression of tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) upon CBX7 knockdown |
GBM: glioblastoma multiforme; HMGA1: high mobility group A1; CCNE: encoding cyclin E
Piles of evidence indicate that elevated expression of CBX proteins (CBX2, CBX7, and CBX8), together with other PRC1 subunits such as BMI1, directly causes transcription repression of the
Ink4a/
Arf locus. The locus encodes two independent tumor suppressors, INK4a and ARF, which activate the retinoblastoma (Rb) and
p53 tumor suppressor pathways, respectively. Regulation of the expression of this locus by CBX protein-containing PRC1 modulates the balance between cell proliferation and senescence (Bernard et al.,
2005; Scott et al.,
2007; Maertens et al.,
2009; Zhang et al.,
2010).
Loss of
Cbx7 expression may be strictly correlated with the acquisition of invasiveness accompanied in a process called the epithelial mesenchymal transition (EMT), which has been seen in several types of cancer, such as thyroid cancer, lung cancer, colon cancer, and pancreatic cancer. CBX7 was found to control the cell cycle and cell proliferation via regulation of the expression of the encoding cyclin E 1 (
CCNE1). CBX7 directly competed with the transcription activator HMGA1 for binding to the promoter of CCNE1, therefore causing down-regulation of the CCNE1. Lack of the CBX7 protein in human lung cancer carcinomas correlates with over-expression of CCNE1 (Forzati et al.,
2012a;
2012b).
CBX7 and EZH2 are also known to physically associate with DNA methyltransferases (DNMTs), responsible for programming the altered DNA methylation profiles observed in multiple cancer types (Mohammad et al.,
2009). This interplay uncovered another mechanism by which multiple sources of epigenetic regulators were integrated to silence their common targets. CBX7 was also known to influence early breast cancer development by controlling microRNA expression (Hannafon et al.,
2011), while in breast cancer, CBX8 cooperated with SIRT1 for suppressing p53 acetylation induced by sirtinol and etoposide/TSA (Lee et al.,
2013). Upon ectopic expression, CBX8 or SIRT1 repressed the expression of p21 by inhibiting p53 binding to the promoter.
The most studied components of PcG complexes in cancer research are EZH2 and BMI1, and they both are often over-expressed in cancers (Mills,
2010). They have been shown to be essential in controlling proliferation and maintaining self-renewal of cancer stem cells in various models (Crea et al.,
2012). Some PRC2 inhibitors, such as DZNeP (a non-specific inhibitor of histone methylation), have been tested on cancer models and exhibited effectiveness in abolishing cancer stem cells self-renewal and tumorigenicity (Crea et al.,
2012). The association between CBX proteins in cancer cells and stem cell proliferation makes them become the focus of ongoing investigations in cancer research. The functional specification among CBX family members provides a reasonable hypothesis that each CBX protein may play very unique roles in various lineages and alter the behavior of normal or transformed stem cells in specific contexts.
7. Concluding remarks
Epigenetic regulation by PcG complexes plays a crucial role in maintaining stemness and determining stem cell fates. The diversity of the composition of PcG complexes, particularly PRC1 has been found to be far more complicated than initially estimated. Recent studies uncovered the function specification of the PRC1 components, such as CBX family members, in fine-tuning the cell fate of the different lineages. These discoveries may help explain the reason why both aberrant up- and down-regulation of PcG proteins have been detected in distinct types of cancer. Further exploration of the specified functions of PcG proteins and the detailed molecular consequences triggered by abnormally acting PcG complexes would be extremely meaningful. At the same time, a lot more remains to be investigated and discovered relating to the interplay among PcG functions and other transcription regulatory mechanisms including other epigenetic regulators, ncRNAs, transcription machinery, and transcription factors. In addition, there is a plethora of post-translational modifications of PcG proteins identified in various cell types and tissues with little known mechanisms. Altogether, increasing knowledge about the roles of PcG proteins in stem cell biology and oncology can help to explore these epigenetic modifiers as potential biomarkers for cancer and further develop therapeutic strategies for early diagnosis, prognosis, and treatment of cancer.
* Project supported by the Fundamental Research Funds for the Central Universities from Lanzhou University (No. lzujbky-2014-87), ChinaAuthors contributions Rong-gang MA, Yang ZHANG, and Ting-ting SUN collected all the references, made all the figures and tables, and wrote a draft. Bo CHENG wrote the manuscript.Compliance with ethics guidelines Rong-gang MA, Yang ZHANG, Ting-ting SUN, and Bo CHENG declare that they have no conflict of interest.References
[1] Agrawal, N., Banerjee, R., 2008. Human polycomb 2 protein is a SUMO E3 ligase and alleviates substrate-induced inhibition of cystathionine β-synthase sumoylation.
PLoS ONE, 3(12):e4032


[2] Aloia, L., Di Stefano, B., Di Croce, L., 2013. Polycomb complexes in stem cells and embryonic development.
Development, 140(12):2525-2534.
[3] Atchison, L., Ghias, A., Wilkinson, F., 2003. Transcription factor YY1 functions as a PcG protein
in vivo
.
EMBO J, 22(6):1347-1358.
[4] Basu, A., Wilkinson, F.H., Colavita, K., 2014. YY1 DNA binding and interaction with YAF2 is essential for polycomb recruitment.
Nucl Acids Res, 42(4):2208-2223.
[5] Baumann, C., de la Fuente, R., 2011. Role of polycomb group protein Cbx2/M33 in meiosis onset and maintenance of chromosome stability in the mammalian germline.
Genes, 2(1):59-80.
[6] Bengani, H., Mendiratta, S., Maini, J., 2013. Identification and validation of a putative polycomb responsive element in the human genome.
PLoS ONE, 8(6):e67217
[7] Bernard, D., Martinez-Leal, J.F., Rizzo, S., 2005. CBX7 controls the growth of normal and tumor-derived prostate cells by repressing the
Ink4a/Arf locus.
Oncogene, 24(36):5543-5551.
[8] Bernstein, B.E., Mikkelsen, T.S., Xie, X., 2006. A bivalent chromatin structure marks key developmental genes in embryonic stem cells.
Cell, 125(2):315-326.
[9] Bernstein, E., Duncan, E.M., Masui, O., 2006. Mouse polycomb proteins bind differentially to methylated histone H3 and RNA and are enriched in facultative heterochromatin.
Mol Cell Biol, 26(7):2560-2569.
[10] Bezsonova, I., Walker, J.R., Bacik, J.P., 2009. Ring1B contains a ubiquitin-like docking module for interaction with Cbx proteins.
Biochemistry, 48(44):10542-10548.
[11] Biason-Lauber, A., Konrad, D., Meyer, M., 2009. Ovaries and female phenotype in a girl with 46, XY karyotype and mutations in the
CBX2 gene.
Am J Hum Genet, 84(5):658-663.
[12] Brockdorff, N., 2013. Noncoding RNA and polycomb recruitment.
RNA, 19(4):429-442.
[13] Brookes, E., de Santiago, I., Hebenstreit, D., 2012. Polycomb associates genome-wide with a specific RNA polymerase II variant, and regulates metabolic genes in ESCs.
Cell Stem Cell, 10(2):157-170.
[14] Brown, J.L., Kassis, J.A., 2013. Architectural and functional diversity of polycomb group response elements in
Drosophila
.
Genetics, 195(2):407-419.
[15] Buchwald, G., van der Stoop, P., Weichenrieder, O., 2006. Structure and E3-ligase activity of the ring-ring complex of polycomb proteins Bmi1 and Ring1b.
EMBO J, 25(11):2465-2474.
[16] Buschbeck, M., Uribesalgo, I., Wibowo, I., 2009. The histone variant macroH2A is an epigenetic regulator of key developmental genes.
Nat Struct Mol Biol, 16(10):1074-1079.
[17] Cantin, G.T., Yi, W., Lu, B., 2008. Combining protein-based IMAC, peptide-based IMAC, and MudPIT for efficient phosphoproteomic analysis.
J Proteome Res, 7(3):1346-1351.
[18] Chen, R.Q., Yang, Q.K., Lu, B.W., 2009. CDC25B mediates rapamycin-induced oncogenic responses in cancer cells.
Cancer Res, 69(6):2663-2668.
[19] Cor, N., Bel, S., Gaunt, S.J., 1997. Altered cellular proliferation and mesoderm patterning in polycomb-M33-deficient mice.
Development, 124(3):721-729.
[20] Cor, N., Joly, F., Boned, A., 2004. Disruption of E2F signaling suppresses the
INK4a-induced proliferative defect in M33-deficient mice.
Oncogene, 23(46):7660-7668.
[21] Crea, F., Paolicchi, E., Marquez, V.E., 2012. Polycomb genes and cancer: time for clinical application?.
Crit Rev Oncol Hematol, 83(2):184-193.
[22] Creyghton, M.P., Markoulaki, S., Levine, S.S., 2008. H2AZ is enriched at polycomb complex target genes in ES cells and is necessary for lineage commitment.
Cell, 135(4):649-661.
[23] Daub, H., Olsen, J.V., Bairlein, M., 2008. Kinase-selective enrichment enables quantitative phospho-proteomics of the kinome across the cell cycle.
Mol Cell, 31(3):438-448.
[24] Dephoure, N., Zhou, C., Villen, J., 2008. A quantitative atlas of mitotic phosphorylation.
PNAS, 105(31):10762-10767.
[25] Dietrich, N., Bracken, A.P., Trinh, E., 2007. Bypass of senescence by the polycomb group protein CBX8 through direct binding to the INK4A-ARF locus.
EMBO J, 26(6):1637-1648.
[26] Dou, H., Huang, C., van Nguyen, T., 2011. SUMOylation and de-SUMOylation in response to DNA damage.
FEBS Lett, 585(18):2891-2896.
[27] Forzati, F., Federico, A., Pallante, P., 2012. CBX7 is a tumor suppressor in mice and humans.
J Clin Invest, 122(2):612-623.
[28] Forzati, F., Federico, A., Pallante, P., 2012. Tumor suppressor activity of CBX7 in lung carcinogenesis.
Cell Cycle, 11(10):1888-1891.
[29] Galanty, Y., Belotserkovskaya, R., Coates, J., 2009. Mammalian SUMO E3-ligases PIAS1 and PIAS4 promote responses to DNA double-strand breaks.
Nature, 462(7275):935-939.
[30] Gao, Z., Zhang, J., Bonasio, R., 2012. PCGF homologs, CBX proteins, and RYBP define functionally distinct PRC1 family complexes.
Mol Cell, 45(3):344-356.
[31] Gil, J., Bernard, D., Martinez, D., 2004. Polycomb CBX7 has a unifying role in cellular lifespan.
Nat Cell Biol, 6(1):67-72.
[32] Hannafon, B.N., Sebastiani, P., de las Morenas, ., 2011. Expression of microRNA and their gene targets are dysregulated in preinvasive breast cancer.
Breast Cancer Res, 13(2):R24
[33] Hatano, A., Matsumoto, M., Higashinakagawa, T., 2010. Phosphorylation of the chromodomain changes the binding specificity of Cbx2 for methylated histone H3.
Biochem Biophys Res Commun, 397(1):93-99.
[34] Hinz, S., Kempkensteffen, C., Christoph, F., 2008. Expression parameters of the polycomb group proteins BMI1, SUZ12, RING1 and CBX7 in urothelial carcinoma of the bladder and their prognostic relevance.
Tumour Biol, 29(5):323-329.
[35] Ismail, I.H., Gagne, J.P., Caron, M.C., 2012. CBX4-mediated SUMO modification regulates BMI1 recruitment at sites of DNA damage.
Nucl Acids Res, 40(12):5497-5510.
[36] Kagey, M.H., Melhuish, T.A., Wotton, D., 2003. The polycomb protein Pc2 is a SUMO E3.
Cell, 113(1):127-137.
[37] Kang, X., Qi, Y., Zuo, Y., 2010. SUMO-specific protease 2 is essential for suppression of polycomb group protein-mediated gene silencing during embryonic development.
Mol Cell, 38(2):191-201.
[38] Kanhere, A., Viiri, K., Araujo, C.C., 2010. Short RNAs are transcribed from repressed polycomb target genes and interact with polycomb repressive complex-2.
Mol Cell, 38(5):675-688.
[39] Karamitopoulou, E., Pallante, P., Zlobec, I., 2010. Loss of the CBX7 protein expression correlates with a more aggressive phenotype in pancreatic cancer.
Eur J Cancer, 46(8):1438-1444.
[40] Katoh-Fukui, Y., Tsuchiya, R., Shiroishi, T., 1998. Male-to-female sex reversal in M33 mutant mice.
Nature, 393(6686):688-692.
[41] Katoh-Fukui, Y., Owaki, A., Toyama, Y., 2005. Mouse polycomb M33 is required for splenic vascular and adrenal gland formation through regulating Ad4BP/SF1 expression.
Blood, 106(5):1612-1620.
[42] Katoh-Fukui, Y., Miyabayashi, K., Komatsu, T., 2012. Cbx2, a polycomb group gene, is required for
Sry gene expression in mice.
Endocrinology, 153(2):913-924.
[43] Klauke, K., Radulovic, V., Broekhuis, M., 2013. Polycomb Cbx family members mediate the balance between haematopoietic stem cell self-renewal and differentiation.
Nat Cell Biol, 15(4):353-362.
[44] Kotake, Y., Nakagawa, T., Kitagawa, K., 2011. Long non-coding RNA ANRIL is required for the PRC2 recruitment to and silencing of
p15INK4B
tumor suppressor gene.
Oncogene, 30(16):1956-1962.
[45] Ku, M., Koche, R.P., Rheinbay, E., 2008. Genomewide analysis of PRC1 and PRC2 occupancy identifies two classes of bivalent domains.
PLoS Genet, 4(10):e1000242
[46] Lee, S.H., Um, S.J., Kim, E.J., 2013. CBX8 suppresses Sirtinol-induced premature senescence in human breast cancer cells via cooperation with SIRT1.
Cancer Lett, 335(2):397-403.
[47] Lee, S.W., Lee, M.H., Park, J.H., 2012. SUMOylation of hnRNP-K is required for p53-mediated cell-cycle arrest in response to DNA damage.
EMBO J, 31(23):4441-4452.
[48] Lewis, E.B., 1978. A gene complex controlling segmentation in
Drosophila
.
Nature, 276(5688):565-570.
[49] Li, B., Zhou, J., Liu, P., 2007. Polycomb protein Cbx4 promotes SUMO modification of
de novo DNA methyltransferase Dnmt3a.
Biochem J, 405(2):369-378.
[50] Li, G., Warden, C., Zou, Z., 2013. Altered expression of polycomb group genes in glioblastoma multiforme.
PLoS ONE, 8(11):e80970
[51] Liu, B., Liu, Y.F., Du, Y.R., 2013. Cbx4 regulates the proliferation of thymic epithelial cells and thymus function.
Development, 140(4):780-788.
[52] Long, J., Zuo, D., Park, M., 2005. Pc2-mediated sumoylation of Smad-interacting protein 1 attenuates transcriptional repression of E-cadherin.
J Biol Chem, 280(42):35477-35489.
[53] Luis, N.M., Morey, L., Mejetta, S., 2011. Regulation of human epidermal stem cell proliferation and senescence requires polycomb-dependent and -independent functions of Cbx4.
Cell Stem Cell, 9(3):233-246.
[54] MacPherson, M.J., Beatty, L.G., Zhou, W., 2009. The CTCF insulator protein is posttranslationally modified by SUMO.
Mol Cell Biol, 29(3):714-725.
[55] Maertens, G.N., El Messaoudi-Aubert, S., Racek, T., 2009. Several distinct polycomb complexes regulate and co-localize on the INK4a tumor suppressor locus.
PLoS ONE, 4(7):e6380
[56] Maethner, E., Garcia-Cuellar, M.P., Breitinger, C., 2013. MLL-ENL inhibits polycomb repressive complex 1 to achieve efficient transformation of hematopoietic cells.
Cell Rep, 3(5):1553-1566.
[57] Malik, B., Hemenway, C.S., 2013. CBX8, a component of the polycomb PRC1 complex, modulates DOT1L-mediated gene expression through AF9/MLLT3.
FEBS Lett, 587(18):3038-3044.
[58] Margueron, R., Li, G., Sarma, K., 2008. Ezh1 and Ezh2 maintain repressive chromatin through different mechanisms.
Mol Cell, 32(4):503-518.
[59] Margueron, R., Justin, N., Ohno, K., 2009. Role of the polycomb protein EED in the propagation of repressive histone marks.
Nature, 461(7265):762-767.
[60] Mendenhall, E.M., Koche, R.P., Truong, T., 2010. GC-rich sequence elements recruit PRC2 in mammalian ES cells.
PLoS Genet, 6(12):e1001244
[61] Merrill, J.C., Kagey, M.H., Melhuish, T.A., 2010. Inhibition of CtBP1 activity by Akt-mediated phosphorylation.
J Mol Biol, 398(5):657-671.
[62] Mills, A.A., 2010. Throwing the cancer switch: reciprocal roles of polycomb and trithorax proteins.
Nat Rev Cancer, 10(10):669-682.
[63] Mohammad, H.P., Cai, Y., McGarvey, K.M., 2009. Polycomb CBX7 promotes initiation of heritable repression of genes frequently silenced with cancer-specific DNA hypermethylation.
Cancer Res, 69(15):6322-6330.
[64] Morey, L., Helin, K., 2010. Polycomb group protein-mediated repression of transcription.
Trends Biochem Sci, 35(6):323-332.
[65] Morey, L., Pascual, G., Cozzuto, L., 2012. Non-overlapping functions of the polycomb group Cbx family of proteins in embryonic stem cells.
Cell Stem Cell, 10(1):47-62.
[66] Morris, J.R., Boutell, C., Keppler, M., 2009. The SUMO modification pathway is involved in the BRCA1 response to genotoxic stress.
Nature, 462(7275):886-890.
[67] Muller, J., Verrijzer, P., 2009. Biochemical mechanisms of gene regulation by polycomb group protein complexes.
Curr Opin Genet Dev, 19(2):150-158.
[68] Muller, J., Gaunt, S., Lawrence, P.A., 1995. Function of the polycomb protein is conserved in mice and flies.
Development, 121(9):2847-2852.
[69] Nacerddine, K., Lehembre, F., Bhaumik, M., 2005. The SUMO pathway is essential for nuclear integrity and chromosome segregation in mice.
Dev Cell, 9(6):769-779.
[70] OLoghlen, A., Munoz-Cabello, A.M., Gaspar-Maia, A., 2012. MicroRNA regulation of Cbx7 mediates a switch of polycomb orthologs during ESC differentiation.
Cell Stem Cell, 10(1):33-46.
[71] Oh, Y., Chung, K.C., 2012. Small ubiquitin-like modifier (SUMO) modification of zinc finger protein 131 potentiates its negative effect on estrogen signaling.
J Biol Chem, 287(21):17517-17529.
[72] Oh, Y., Kim, Y.M., Mouradian, M.M., 2011. Human polycomb protein 2 promotes α-synuclein aggregate formation through covalent SUMOylation.
Brain Res, 1381:78-89.
[73] Oktaba, K., Gutierrez, L., Gagneur, J., 2008. Dynamic regulation by polycomb group protein complexes controls pattern formation and the cell cycle in
Drosophila
.
Dev Cell, 15(6):877-889.
[74] Olsen, J.V., Blagoev, B., Gnad, F., 2006. Global,
in vivo, and site-specific phosphorylation dynamics in signaling networks.
Cell, 127(3):635-648.
[75] Pallante, P., Federico, A., Berlingieri, M.T., 2008. Loss of the
CBX7 gene expression correlates with a highly malignant phenotype in thyroid cancer.
Cancer Res, 68(16):6770-6778.
[76] Pallante, P., Terracciano, L., Carafa, V., 2010. The loss of the
CBX7 gene expression represents an adverse prognostic marker for survival of colon carcinoma patients.
Eur J Cancer, 46(12):2304-2313.
[77] Pasini, D., Bracken, A.P., Jensen, M.R., 2004. Suz12 is essential for mouse development and for EZH2 histone methyltransferase activity.
EMBO J, 23(20):4061-4071.
[78] Pelisch, F., Pozzi, B., Risso, G., 2012. DNA damage-induced heterogeneous nuclear ribonucleoprotein K sumoylation regulates p53 transcriptional activation.
J Biol Chem, 287(36):30789-30799.
[79] Pemberton, H., Anderton, E., Patel, H., 2014. Genome-wide co-localization of polycomb orthologs and their effects on gene expression in human fibroblasts.
Genome Biol, 15(2):R23
[80] Plath, K., Fang, J., Mlynarczyk-Evans, S.K., 2003. Role of histone H3 lysine 27 methylation in X inactivation.
Science, 300(5616):131-135.
[81] Ren, X., Kerppola, T.K., 2011. REST interacts with Cbx proteins and regulates polycomb repressive complex 1 occupancy at RE1 elements.
Mol Cell Biol, 31(10):2100-2110.
[82] Ren, X., Vincenz, C., Kerppola, T.K., 2008. Changes in the distributions and dynamics of polycomb repressive complexes during embryonic stem cell differentiation.
Mol Cell Biol, 28(9):2884-2895.
[83] Richly, H., Aloia, L., di Croce, L., 2011. Roles of the polycomb group proteins in stem cells and cancer.
Cell Death Dis, 2(9):e204
[84] Rikova, K., Guo, A., Zeng, Q., 2007. Global survey of phosphotyrosine signaling identifies oncogenic kinases in lung cancer.
Cell, 131(6):1190-1203.
[85] Roscic, A., Moller, A., Calzado, M.A., 2006. Phosphorylation-dependent control of Pc2 SUMO E3 ligase activity by its substrate protein HIPK2.
Mol Cell, 24(1):77-89.
[86] Schorderet, P., Duboule, D., 2011. Structural and functional differences in the long non-coding RNA hotair in mouse and human.
PLoS Genet, 7(5):e1002071
[87] Schorderet, P., Lonfat, N., Darbellay, F., 2013. A genetic approach to the recruitment of PRC2 at the
HoxD locus.
PLoS Genet, 9(11):e1003951
[88] Schwartz, Y.B., Pirrotta, V., 2013. A new world of polycombs: unexpected partnerships and emerging functions.
Nat Rev Genet, 14(12):853-864.
[89] Scott, C.L., Gil, J., Hernando, E., 2007. Role of the chromobox protein CBX7 in lymphomagenesis.
PNAS, 104(13):5389-5394.
[90] Senthilkumar, R., Mishra, R.K., 2009. Novel motifs distinguish multiple homologues of polycomb in vertebrates: expansion and diversification of the epigenetic toolkit.
BMC Genom, 10:549
[91] Shinjo, K., Yamashita, Y., Yamamoto, E., 2013. Expression of chromobox homolog 7 (CBX7) is associated with poor prognosis in ovarian clear cell adenocarcinoma via TRAIL-induced apoptotic pathway regulation.
Int J Cancer, in press,:
[92] Simon, J., Chiang, A., Bender, W., 1993. Elements of the
Drosophila bithorax complex that mediate repression by polycomb group products.
Dev Biol, 158(1):131-144.
[93] Simon, J.A., Kingston, R.E., 2013. Occupying chromatin: polycomb mechanisms for getting to genomic targets, stopping transcriptional traffic, and staying put.
Mol Cell, 49(5):808-824.
[94] Song, J., Durrin, L.K., Wilkinson, T.A., 2004. Identification of a SUMO-binding motif that recognizes SUMO-modified proteins.
PNAS, 101(40):14373-14378.
[95] Srinivasan, L., Atchison, M.L., 2004. YY1 DNA binding and PcG recruitment requires CtBP.
Genes Dev, 18(21):2596-2601.
[96] Struhl, G., 1981. A gene product required for correct initiation of segmental determination in
Drosophila
.
Nature, 293(5827):36-41.
[97] Tan, J., Jones, M., Koseki, H., 2011. CBX8, a polycomb group protein, is essential for MLL-AF9-induced leukemogenesis.
Cancer Cell, 20(5):563-575.
[98] Tavares, L., Dimitrova, E., Oxley, D., 2012. RYBP-PRC1 complexes mediate H2A ubiquitylation at polycomb target sites independently of PRC2 and H3K27me
3
.
Cell, 148(4):664-678.
[99] Tsai, M.C., Manor, O., Wan, Y., 2010. Long noncoding RNA as modular scaffold of histone modification complexes.
Science, 329(5992):689-693.
[100] van den Boom, V., Rozenveld-Geugien, M., Bonardi, F., 2013. Nonredundant and locus-specific gene repression functions of PRC1 paralog family members in human hematopoietic stem/progenitor cells.
Blood, 121(13):2452-2461.
[101] Vincenz, C., Kerppola, T.K., 2008. Different polycomb group CBX family proteins associate with distinct regions of chromatin using nonhomologous protein sequences.
PNAS, 105(43):16572-16577.
[102] Wang, B., Tang, J., Liao, D., 2013. Chromobox homolog 4 is correlated with prognosis and tumor cell growth in hepatocellular carcinoma.
Ann Surg Oncol, 20(S3):S684-S692.
[103] Wang, H., Wang, L., Erdjument-Bromage, H., 2004. Role of histone H2A ubiquitination in polycomb silencing.
Nature, 431(7010):873-878.
[104] Woo, C.J., Kharchenko, P.V., Daheron, L., 2010. A region of the human HOXD cluster that confers polycomb-group responsiveness.
Cell, 140(1):99-110.
[105] Woo, C.J., Kharchenko, P.V., Daheron, L., 2013. Variable requirements for DNA-binding proteins at polycomb-dependent repressive regions in human HOX clusters.
Mol Cell Biol, 33(16):3274-3285.
[106] Wotton, D., Merrill, J.C., 2007. Pc2 and SUMOylation.
Biochem Soc Trans, 35(Pt 6):1401-1404.
[107] Wu, H.A., Balsbaugh, J.L., Chandler, H., 2013. Mitogen-activated protein kinase signaling mediates phosphorylation of polycomb ortholog
Cbx7
.
J Biol Chem, 288(51):36398-36408.
[108] Wu, X., Johansen, J.V., Helin, K., 2013. Fbxl10/Kdm2b recruits polycomb repressive complex 1 to CpG islands and regulates H2A ubiquitylation.
Mol Cell, 49(6):1134-1146.
[109] Yang, L., Lin, C., Liu, W., 2011. ncRNA- and Pc2 methylation-dependent gene relocation between nuclear structures mediates gene activation programs.
Cell, 147(4):773-788.
[110] Yap, K.L., Li, S., Munoz-Cabello, A.M., 2010. Molecular interplay of the noncoding RNA ANRIL and methylated histone H3 lysine 27 by polycomb CBX7 in transcriptional silencing of
INK4a
.
Mol Cell, 38(5):662-674.
[111] Zhang, X.W., Zhang, L., Qin, W., 2010. Oncogenic role of the chromobox protein CBX7 in gastric cancer.
J Exp Clin Cancer Res, 29(1):114
[112] Zhao, J., Sun, B.K., Erwin, J.A., 2008. Polycomb proteins targeted by a short repeat RNA to the mouse X chromosome.
Science, 322(5902):750-756.
[113] Zhao, J., Ohsumi, T.K., Kung, J.T., 2010. Genome-wide identification of polycomb-associated RNAs by RIP-seq.
Mol Cell, 40(6):939-953.
[114] Zhou, X., Zhang, H.L., Gu, G.F., 2013. Investigation of the relationship between chromobox homolog 8 and nucleus pulposus cells degeneration in rat intervertebral disc.
In Vitro Cell Dev Biol Anim, 49(4):279-286.



Open peer comments: Debate/Discuss/Question/Opinion
<1>