1. Introduction
Dioscorea bulbifera L. (Dioscoreaceae) rhizome is widely distributed throughout the tropics and temperate regions, especially in Asia, and in China, it has been traditionally used to treat thyroid disease and tumors (Gao et al.,
2002). In addition, in northern Bangladesh, it is used to treat tumors and leprosy (Murray et al.,
1984). Our laboratory studies have shown that
D. bulbifera rhizome can induce hepatotoxicity (Wang et al.,
2010;
2011), which emphasizes a potential toxicity in the clinical setting. Among the biologically active substances in
D. bulbifera rhizome, a major class is the clerodane diterpenoids (Wang et al.,
2009; Liu et al.,
2010). Diosbulbin B is the major chemical compound of the clerodane diterpenoids in
D. bulbifera rhizome (Kawasaki et al.,
1968), and has been found to have anti-tumor activity (Gao et al.,
2002; Wang et al.,
2012).
Angelica sinensis (Oliv.) Diels root has been reported to prevent the hepatotoxicity induced by
D. bulbifera rhizome (Liu et al.,
2004), while the active compound has not yet been elucidated. Ferulic acid is one of the main bioactive molecules in
A. sinensis root, and always acts as a strong anti-oxidant; it is known for treating skin cancer, ageing, fatigue, and muscle wasting (Graf,
1992; Alias et al.,
2009; Barone et al.,
2009; You et al.,
2009). There are reports that ferulic acid prevents noise-induced hearing loss and free radical-induced skin damage (Calabrese et al.,
2008; Fetoni et al.,
2010), but whether it affects the hepatotoxicity and anti-tumor activity of diosbulbin B is still unknown.
Various reports have demonstrated that oxidative stress plays a vital role in liver toxicity induced by such substances as alcohol, carbon tetrachloride, chemotherapeutic agents, and acetaminophen (dos Santos et al.,
2007; Kostopanagiotou et al.,
2009; Moselhy and Ali,
2009; Samuhasaneeto et al.,
2009). Reactive oxygen species (ROS), generated during oxidative stress, are extremely reactive and may modify and inactivate cellular lipids, proteins, DNA, and RNA, and thus cause cell damage. The cellular anti-oxidant system in the body prevents ROS-induced cell injury; the system includes low-molecular-mass anti-oxidants (van der Vliet et al.,
1999) such as glutathione, α-tocopherol, ascorbic acid, and the main anti-oxidant enzymes including copper- and zinc-containing superoxide dismutase (CuZn-SOD) and catalase (CAT). Our previous report demonstrated that oxidative stress plays an important role in liver injury induced by acetate extracts of
D. bulbifera (Wang et al.,
2011); however, whether the protection by ferulic acid against liver injury induced by diosbulbin B is related to the liver anti-oxidant system has not been demonstrated.
The present study is designed to demonstrate the protection by ferulic acid against the hepatotoxicity induced by diosbulbin B and the underlying mechanism, and investigate whether ferulic acid enhances diosbulbin B-inhibited tumor growth.
2. Materials and methods
2.1. Experimental animals
ICR male mice (18–22 g) were purchased from the Shanghai Laboratory Animal Center of Chinese Academy of Sciences, China. Animals were given rodent laboratory chow and water
ad libitum, and maintained under controlled conditions at (22±1) °C, relative humidity (65±10)%, and a 12 h/12 h light/dark cycle (lights on at 7:00 am). All animals were dealt with in compliance with the institutional animal care guidelines of the Experimental Animal Ethical Committee of Shanghai University of Traditional Chinese Medicine, China.
2.2. Cell lines
Mouse sarcoma S180 tumor cells were maintained in the peritoneal cavities of ICR mice in the Laboratory of Experimental Oncology of the Shanghai Laboratory Animal Center of Chinese Academy of Sciences, China.
2.3. Reagents
5-Fluorouracil was purchased from Shanghai Bangcheng Chemical Co., Ltd., China. Unless indicated, other reagents were purchased from Sigma Chemical Co. (St. Louis, MO, USA).
2.4. Preparations of diosbulbin B and ferulic acid
D. bulbifera rhizome was collected in Qingyang County, Anhui Province, China and authenticated by Prof. Shou-jin LIU (Anhui University of Chinese Medicine, China).
A. sinensis root was purchased from Minxian County, Gansu Province, China and authenticated by Prof. Li-hong WU (Shanghai University of Traditional Chinese Medicine, China). The voucher specimens of
D. bulbifera rhizome and
A. sinensis root were both deposited in the herbarium of the Institute of Traditional Chinese Medicine, Shanghai University of Traditional Chinese Medicine, China.
Diosbulbin B and ferulic acid were isolated from
D. bulbifera rhizome and
A. sinensis root, respectively, in our laboratory as previously described (Kawasaki et al.,
1968; Sun et al.,
2006). After purification using silica gel column and gel chromatography, purities of diosbulbin B and ferulic acid were both more than 98% as determined by high-performance liquid chromatography (HPLC) with diode array detector. The chemical structures of diosbulbin B and ferulic acid are shown in Fig.
1.
Fig.1
Chemical structures of diosbulbin B (a) and ferulic acid (b)
2.5. Treatment protocol
Ascites of the mouse sarcoma S180 were drawn out from mice under aseptic conditions. Cells were counted promptly and then diluted to about 1×10
7 cells/ml with aseptic saline. The diluted S180 cell suspension was inoculated subcutaneously (0.1 ml per mouse) into the right armpit (Bezerra et al.,
2008).
One day after inoculation, mice, except for the normal (non-tumor-inoculated) animals, were randomly divided into five groups of eight mice each. 5-Fluorouracil-treated mice were injected intraperitoneally with 5-fluorouracil at a dose of 25 mg/kg once every other day. Normal (non-tumor-inoculated) and control (tumor-inoculated) groups of mice received daily oral administration of 0.5% (5 g/L) sodium carboxyl methyl cellulose (CMC-Na; 0.2 ml/10 g). The other three groups received ferulic acid (8 mg/kg), diosbulbin B (32 mg/kg), or diosbulbin B (32 mg/kg) plus ferulic acid (8 mg/kg) by intragastric administration for 12 d starting from 24 h after tumor inoculation. After treatment, mice were sacrificed by cervical dislocation after peripheral blood samples, liver tissues and tumors were collected at 24 h after the last administration. Serum samples were collected for the determination of alanine transaminase/aspartate transaminase (ALT/AST), and liver tissues were used for the analysis of the lipid peroxide level, histological observation, determination of glutathione-related and anti-oxidant enzymes, and mRNA expression of the main anti-oxidant enzymes. The tumors were weighed, arrayed in line on paper, and taken pictures. The tumor inhibition ratio (IR) was calculated by the formula of IR=[(
C−
T)/
C]×100%, where
C and
T are the mean tumor weights of the control group (CMC-Na) and the treated group, respectively.
2.6. Assay for serum ALT and AST
Blood samples were obtained from mice of all groups (eight mice per group) for the determination of ALT and AST. Serum ALT and AST were assayed according to Kamei et al. (
1986).
2.7. Assay for liver lipid peroxide level
Liver tissues were homogenized in cold phosphate buffered saline (PBS). Lipid peroxide was determined as previously described (Nkosi et al.,
2006). Malondialdehyde is an end product of lipid peroxide and serves as a means of quantifying lipid peroxide. Malondialdehyde reacts with 2-thiobarbituric acid to generate a pink-colored product, which has an absorbance at 532 nm. The standard curve of malondialdehyde was constructed over the concentration range of 0–40 μmol/L. Lipid peroxide level was expressed as micromoles of malondialdehyde per milligram of protein based on tissue protein concentration.
2.8. Histological observation
After fixation in formaldehyde, the livers were examined for size, color changes, and hemorrhage. Slices of liver were cut into small pieces and histological sections were stained with hematoxylin and eosin (H&E) for the observation under light microscopy.
2.9. Enzymatic assays
Tissues were homogenized in cold PBS, and then centrifuged at 5000×
g for 5 min and the supernatant transferred to new tubes for assay. The liver tissue activities of CuZn-SOD and CAT were determined as previously described (Marklund and Marklund,
1974; Aebi,
1984), and the results were all calculated based on tissue protein concentrations.
2.10. Reverse transcription-polymerase chain reaction (RT-PCR) analysis
Total RNA was extracted from the liver tissue using Trizol (Life Technologies, USA) according to the manufacturer’s protocol. To synthesize single-strand cDNA, reverse transcription of 2 μg total RNA was carried out using 200 U Moloney murine leukemia virus reverse transcriptase (M-MLV RT) and reaction mixture (2.5 mmol/L dNTP, 100 pmol/L oligo(dT) primer, RT buffer, 50 U RNA inhibitor) according to the manufacturer’s protocol. Transcripts of the gene for glyceraldehyde-3-phosphate dehydrogenase (GAPDH) were used as an internal control. Sequences of the PCR primers were as follows: CuZn-SOD forward 5'-AAG GCC GTG TGC GTG CTG AA-3', reverse 5'-CAG GTC TCC AAC ATG CCT CT-3' (246 bp product) (El Mouatassim et al.,
1999); CAT forward 5'-GCA GAT ACC TGT GAA CTG TC-3', reverse 5'-GTA GAA TGT CCG CAC CTG AG-3' (229 bp product) (El Mouatassim et al.,
1999); and, GAPDH forward 5'-GAC CCC TTC ATT GAC CTC AAC T-3', Reverse 5'-GTT TGT GAT GGG TGT GAA CCA-3' (200 bp product) (Hougardy et al.,
2005). The PCR protocol consisted of denaturation at 94 °C for 1 min, 35 cycles of denaturation at 94 °C for 45 s, annealing at 56 °C (CuZn-SOD) or 55 °C (CAT) for 1 min, extension at 72 °C for 1 min, and final extension at 72 °C for 15 min. The protocol for GAPDH was the same except for annealing at 65 °C for 1 min and amplification for 25 cycles. The PCR products were electrophoresed in a 2% agarose gel and stained with ethidium bromide. The bands were automatically analyzed by Smart View Bio-electrophoresis Image Analysis system (Version FR-980, FURI Science and Technology Co. Ltd., Shanghai, China).
2.11. Statistical analysis
All experimental data were expressed as mean±standard error of mean (SEM). Significant differences among experimental groups were compared by one-way analysis of variance (ANOVA) followed by least significant difference (LSD) (
P<0.05) using the Statistics Package for Social Science (SPSS) program Version 11.5.
3. Results
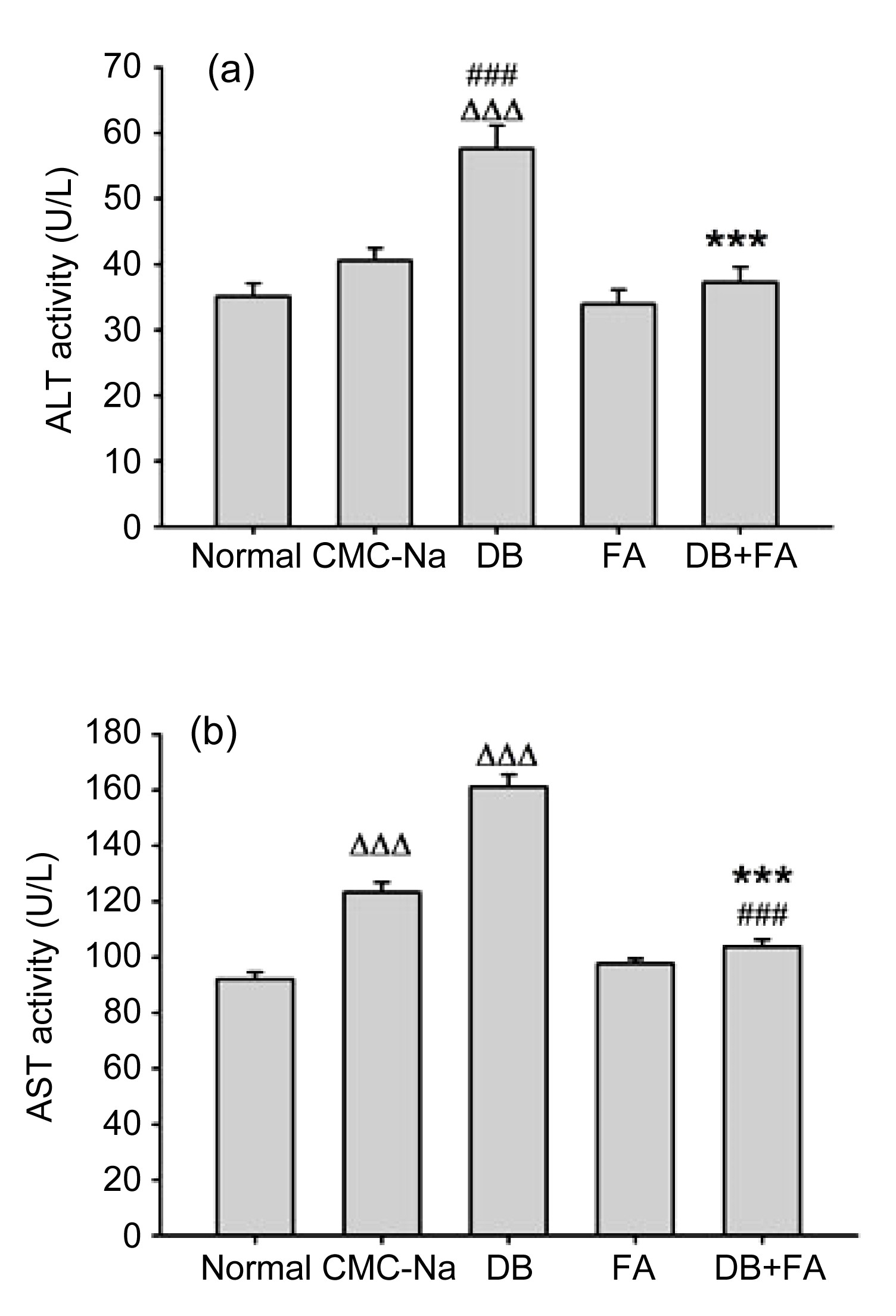
3.1. Ferulic acid prevents diosbulbin B-induced liver injury
Serum ALT and AST activities are liver injury biomarkers, a significant elevation of which often reflects liver injury (Kamei et al.,
1986). In the present study, ALT and AST were both found to be raised in mice treated with diosbulbin B alone for 12 consecutive days compared with CMC-Na controls (
P<0.001), while ferulic acid inhibited this diosbulbin B-induced increase (
P<0.001) (Fig.
2).
Fig.2
Effect of ferulic acid (FA) on ALT (a) and AST (b) activities
FA inhibited diosbulbin B (DB)-induced increase of ALT/AST in tumor-bearing mice. Data are presented as mean±SEM (n=8). ∆∆∆
P<0.001 vs. normal; ###
P<0.001 vs. CMC-Na; ***
P<0.001 vs. DB
As shown in Fig.
3, malondialdehyde, one of the main end-products of lipid peroxide activity (Nkosi et al.,
2006), was increased in the livers of mice treated with diosbulbin B compared with CMC-Na controls (
P<0.05), while ferulic acid inhibited this increase (
P<0.001).
Fig.3
Effect of ferulic acid (FA) on malondialdehyde (MDA) level
FA inhibited diosbulbin B (DB)-increased MDA amount. Data are presented as mean±SEM (n=8). ∆∆
P<0.01 vs. normal; #
P<0.05 vs. CMC-Na; ***
P<0.001 vs. DB
Further, histological evaluation of the livers removed from mice treated with diosbulbin B (32 mg/kg) showed hydropic degeneration of hepatocytes (Fig.
4c). After treatment with ferulic acid (8 mg/kg), these abnormal changes disappeared (Fig.
4d).
Fig.4
Liver histology of the normal (a), CMC-Na control (b), diosbulbin B (32 mg/kg) (c), and diosbulbin B (32 mg/kg)+ferulic acid (8 mg/kg) (d) groups
After treatment, livers were removed, fixed, sectioned (5 μm), and processed for H&E staining. Typical images were chosen from each experimental group (original magnification 200×)
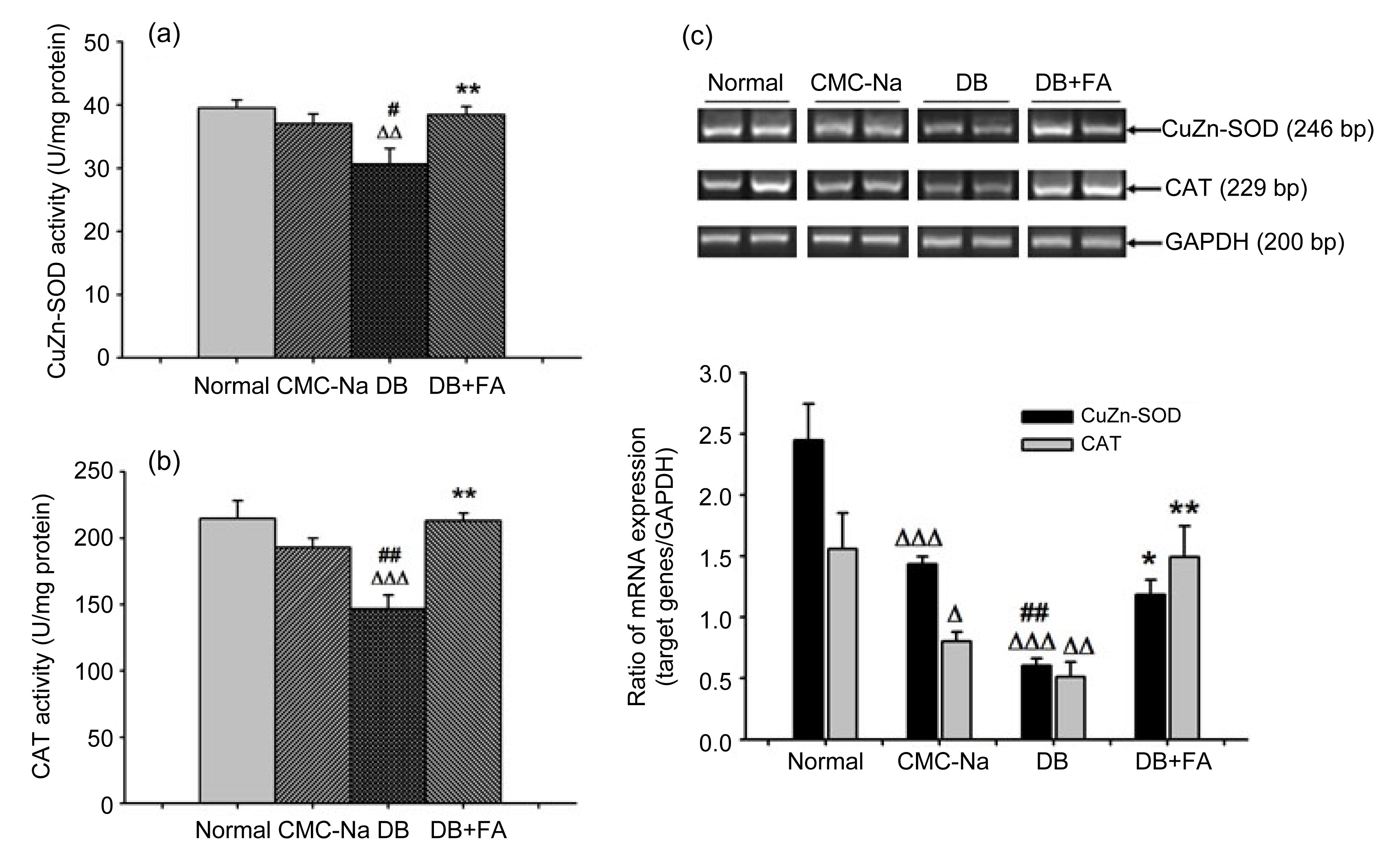
3.2. Ferulic acid reverses diosbulbin B-decreased CuZn-SOD and CAT enzymatic activities and mRNA expression
CAT and CuZn-SOD are both important intracellular anti-oxidant enzymes, participating in the process of oxidative stress (Aebi,
1984; Zelko et al.,
2002). Our results showed that diosbulbin B decreased the enzymatic activities of CuZn-SOD and CAT compared with CMC-Na controls (
P<0.05 and
P<0.01, respectively), while ferulic acid reversed this decrease (both
P<0.01) (Figs.
5a and 5b). Further results (Fig.
5c) showed that in the CMC-Na control group, mRNA expressions of CAT and CuZn-SOD were decreased compared with the normal (
P<0.05 and
P<0.001, respectively). The mRNA expressions of CAT and CuZn-SOD in diosbulbin B-treated mice were further decreased compared with the CMC-Na group (both
P<0.01), while ferulic acid clearly reversed this decrease (
P<0.05 and
P<0.01, respectively).
Fig.5
Effects of ferulic acid (FA) on CuZn-SOD and CAT enzymatic activity and mRNA expression
FA reversed diosbulbin B (DB)-decreased liver CuZn-SOD (a) and CAT (b) enzymatic activity and mRNA expression (c). Data are presented as mean±SEM, with n=8 (a, b) or n=6 (c). ∆
P<0.05, ∆∆
P<0.01, ∆∆∆
P<0.001 vs. normal; #
P<0.05, ##
P<0.01 vs. CMC-Na; *
P<0.05, **
P<0.01 vs. DB
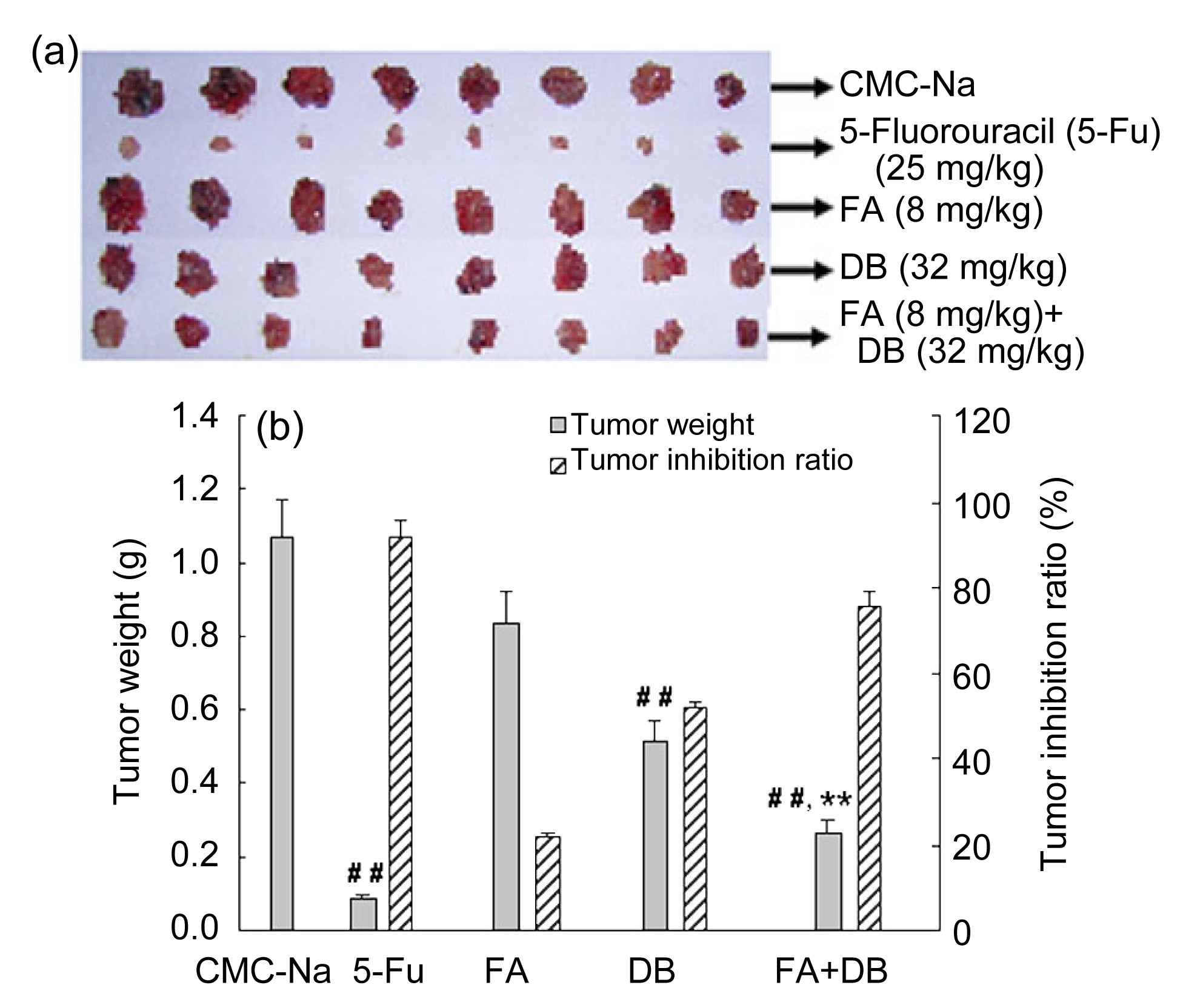
3.3. Ferulic acid enhances diosbulbin B-induced anti-tumor activity
The effect of ferulic acid on the anti-tumor activity of diosbulbin B in transplanted S180 sarcoma is shown in Fig.
6. Our results showed that diosbulbin B decreased tumor weight of tumor-bearing mice (
P<0.01) by 51.8%. After treatment with ferulic acid, diosbulbin B-decreased tumor weight was further decreased by 75.5% compared with diosbulbin B alone (
P<0.01). Meanwhile, ferulic acid itself had no demonstrable inhibition of tumor growth.
Fig.6
Effect of ferulic acid (FA) on anti-tumor activity
FA augmented diosbulbin B (DB)-induced anti-tumor activity in vivo. After treatment, the tumors were excised and weighed. (a) Pictures of tumor; (b) Tumor weight and the inhibitory ratio of tumor growth. Data are presented as mean±SEM (n=8). ##
P<0.01 vs. CMC-Na; **
P<0.01 vs. DB
4. Discussion
Much attention has been paid to the prevention of cancer (Glade,
1999). In addition to western therapies for cancer prevention, there are also studies on anti-tumor effects of herbal medicines such as
D. bulbifera,
Tripterygium wilfordii, and
Andrographis paniculata (Shamon et al.,
1997; Gao et al.,
2002; Kumar et al.,
2004). Compatibility is a main principle of Chinese medicinal herbs, and some reports have demonstrated that compatibility may lead to eliminating toxicity or reinforcing bioactivities (Kim et al.,
2007; Gao et al.,
2009). Ferulic acid is the main bioactive component of
A. sinensis root, and our results from ALT/AST assays and histological observations are the first to demonstrate that ferulic acid prevents diosbulbin B-induced liver injury. Furthermore, the results show that ferulic acid enhances the diosbulbin B-induced anti-tumor effect. All these results indicate that the compatibility of
A. sinensis root with
D. bulbifera may relieve the hepatotoxicity caused by
D. bulbifera, and provide the possibility of clinical applications of these two drugs for cancer therapy.
The imbalance of oxidants and anti-oxidants will lead to oxidative stress in the liver. Further, many anti-oxidant enzymes and non-enzymatic anti-oxidants may be changed during this process (Łuczaj and Skrzydlewska,
2004; Dadkhah et al.,
2006; Limón-Pacheco and Gonsebatt,
2009). Among them, lipid peroxide is free radical-related (Romero et al.,
1998), and one of the main end-products of lipid peroxide is malondialdehyde. The results show that ferulic acid inhibited the diosbulbin B-induced increase in malondialdehyde, indicating that ferulic acid can prevent diosbulbin B-induced lipid peroxide injury in the liver.
CAT and SOD are believed to play key roles in the enzymatic defense of cells against oxidative stress injury. CAT mainly exists in the peroxisomes of aerobic cells and serves to protect against damage from hydrogen peroxide by catalyzing it into molecular oxygen and water without producing toxic free radicals (Bocchetti and Regoli,
2006; Yilmaz et al.,
2006). The metalloenzyme, SOD, can convert two molecules of superoxide, produced during the oxidative stress, to hydrogen peroxide (Bocchetti and Regoli,
2006). There are three SOD isoenzymes in mammalian cells: CuZn-SOD (copper- and zinc-containing SOD, mainly cytosolic), Mn SOD (manganese-containing SOD, located in the mitochondria), and EC SOD (extracellular SOD, actually also CuZn-SOD) (Zelko et al.,
2002); of these, CuZn-SOD is probably the most important anti-oxidant enzyme (Peskin,
1997). Our results showed that ferulic acid reversed diosbulbin B-decreased enzymatic activities and mRNA expressions of CAT and CuZn-SOD. The results suggest that ferulic acid can prevent diosbulbin B-induced oxidative stress injury, while CAT and CuZn-SOD may also participate in this protection.
In conclusion, the present study shows that ferulic acid can prevent hepatotoxicity induced by diosbulbin B isolated from
D. bulbifera rhizome, mainly via preventing liver oxidative stress injury. This study also demonstrates that ferulic acid can enhance the anti-tumor activity of diosbulbin B in mice. Further studies are in progress in our laboratory to explore the molecular mechanisms of this reinforced anti-tumor activity.
* Project supported by the Program for New Century Excellent Talents in University (No. NCET-11-1054), the National Natural Science Foundation of China (No. 81322053), and the Program for Changjiang Scholars and Innovative Research Team in University (No. PCSIRT-IRT1071), ChinaCompliance with ethics guidelines Jun-ming WANG, Yu-chen SHENG, Li-li JI, and Zheng-tao WANG declare that they have no conflict of interest.References
[1] Aebi, H., 1984. Catalase
in vitro
.
Methods Enzymol, 105:121-126.


[2] Alias, L.M., Manoharan, S., Vellaichamy, L., 2009. Protective effect of ferulic acid on 7,12-dimethylbenz[a] anthracene-induced skin carcinogenesis in Swiss albino mice.
Exp Toxicol Pathol, 61(3):205-214.
[3] Barone, E., Calabrese, V., Mancuso, C., 2009. Ferulic acid and its therapeutic potential as a hormetin for age-related diseases.
Biogerontology, 10(2):97-108.
[4] Bezerra, D.P., Pessoa, C., Moraes, M.O., 2008.
In vivo growth-inhibition of sarcoma 180 by piperlonguminine, an alkaloid amide from the
Piper species.
J Appl Toxicol, 28(5):599-607.
[5] Bocchetti, R., Regoli, F., 2006. Seasonal variability of oxidative biomarkers, lysosomal parameters, metallothioneins and peroxisomal enzymes in the Mediterranean mussel
Mytilus galloprovincialis from Adriatic Sea.
Chemosphere, 65(6):913-921.
[6] Calabrese, V., Calafato, S., Puleo, E., 2008. Redox regulation of cellular stress response by ferulic acid ethyl ester in human dermal fibroblasts: role of vitagenes.
Clin Dermatol, 26(4):358-363.
[7] Dadkhah, A., Fatemi, F., Kazemnejad, S., 2006. Differential effects of acetaminophen on enzymatic and non-enzymatic antioxidant factors and plasma total antioxidant capacity in developing and adult rats.
Mol Cell Biochem, 281(1-2):145-152.
[8] dos Santos, N.A., Martins, N.M., Curti, C., 2007. Dimethylthiourea protects against mitochondrial oxidative damage induced by cisplatin in liver of rats.
Chem Biol Inter, 170(3):177-186.
[9] El Mouatassim, S., Gurin, P., Mnzo, Y., 1999. Expression of genes encoding antioxidant enzymes in human and mouse oocytes during the final stages of maturation.
Mol Hum Reprod, 5(8):720-725.
[10] Fetoni, A.R., Mancuso, C., Eramo, S.L., 2010.
In vivo protective effect of ferulic acid against noise-induced hearing loss in the guinea-pig.
Neuroscience, 169(4):1575-1588.
[11] Gao, H., Kuroyanagi, M., Wu, L., 2002. Antitumor-promoting constituents from
Dioscorea bulbifera L. in JB6 mouse epidermal cells.
Biol Pharm Bull, 25(9):1241-1243.
[12] Gao, J.L., He, T.C., Li, Y.B., 2009. A traditional Chinese medicine formulation consisting of
Rhizoma corydalis and
Rhizoma curcumae exerts synergistic anti-tumor activity.
Oncol Rep, 22(5):1077-1083.
[13] Glade, M.J., 1999. Food, nutrition, and the prevention of cancer: a global perspective (American Institute for Cancer Research/World Cancer Research Fund, American Institute for Cancer Research, 1997).
Nutrition, 15(6):523-526.
[14] Graf, E., 1992. Antioxidant potential of ferulic acid.
Free Radical Biol Med, 13(4):435-448.
[15] Hougardy, B.M., van der Zee, A.G., van den Heuvel, F.A., 2005. Sensitivity to Fas-mediated apoptosis in high-risk HPV-positive human cervical cancer cells: relationship with Fas, caspase-8, and Bid.
Gynecol Oncol, 97(2):353-364.
[16] Kamei, T., Asano, K., Nakamura, S., 1986. Determination of serum glutamate oxaloacetate transaminase and glutamate pyruvate transaminase by using L-glutamate oxidase.
Chem Pharm Bull, 34(1):409-412.
[17] Kawasaki, T., Komori, T., Setoguchi, S., 1968. Furanoid norditerpenes from
Dioscoreacae plants. 1. Diosbulins A, B, and C from
Dioscorea bulbifera form a spontanea.
Chem Pharm Bull, 16(12):2430-2435.
[18] Kim, K.C., Kim, J.S., Son, J.K., 2007. Enhanced induction of mitochondrial damage and apoptosis in human leukemia HL-60 cells by the
Ganoderma lucidum and
Duchesnea chrysantha extracts.
Cancer Lett, 246(1-2):210-217.
[19] Kostopanagiotou, G.G., Grypioti, A.D., Matsota, P., 2009. Acetaminophen-induced liver injury and oxidative stress: protective effect of propofol.
Eur J Anaesthesiol, 26(7):548-553.
[20] Kumar, R.A., Sridevi, K., Kumar, N.V., 2004. Anticancer and immunostimulatory compounds from
Andrographis paniculata
.
J Ethnopharmacol, 92(2-3):291-295.
[21] Limn-Pacheco, J., Gonsebatt, M.E., 2009. The role of antioxidants and antioxidant-related enzymes in protective responses to environmentally induced oxidative stress.
Mutat Res/Genet Toxicol Environ Mutagen, 674(1-2):137-147.
[22] Liu, H., Chou, G.X., Guo, Y.L., 2010. Norclerodane diterpenoids from rhizomes of
Dioscorea bulbifera
.
Phytochemistry, 71(10):1174-1180.
[23] Liu, S.M., Li, Y.J., Luo, M.M., 2004. The detoxification action of Chinese
Angelica root to
Dioscorea bulbifera L.
Chin J Integr Trad West Med Liver Dis, (in Chinese),14(4):216-218.
[24] Łuczaj, W., Skrzydlewska, E., 2004. Antioxidant properties of black tea in alcohol intoxication.
Food Chem Toxicol, 42(12):2045-2051.
[25] Marklund, S.L., Marklund, G., 1974. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase.
Eur J Biochem, 47(3):469-474.
[26] Moselhy, S.S., Ali, H.K., 2009. Hepatoprotective effect of cinnamon extracts against carbon tetrachloride induced oxidative stress and liver injury in rats.
Biol Res, 42(1):93-98.
[27] Murray, R.D.H., Jorge, Z., Khan, N.H., 1984. Diosbulbin D and 8-epidiosbulbin E acetate, norclerodane diterpenoids from
Dioscorea bulbifera tubers.
Phytochemistry, 23(3):623-625.
[28] Nkosi, C.Z., Opoku, A.R., Terblanche, S.E., 2006. Antioxidative effects of pumpkin seed (
Cucurbita pepo) protein isolate in CCl
4-induced liver injury in low-protein fed rats.
Phytother Res, 20(11):935-940.
[29] Peskin, A.V., 1997. Cu, Zn-superoxide dismutase gene dosage and cell resistance to oxidative stress: a review.
Biosci Rep, 17(1):85-89.
[30] Romero, F.J., Bosch-Morell, F., Romero, M.J., 1998. Lipid peroxidation products and antioxidants in human disease.
Environ Health Perspect, 106(Suppl. 5):1229-1234.
[31] Samuhasaneeto, S., Thong-Ngam, D., Kulaputana, O., 2009. Curcumin decreased oxidative stress, inhibited NF-κB activation, and improved liver pathology in ethanol-induced liver injury in rats.
J Biomed Biotechnol, 2009:981963
[32] Shamon, L.A., Pezzuto, J.M., Graves, J.M., 1997. Evaluation of the mutagenic, cytotoxic, and antitumor potential of triptolide, a highly oxygenated diterpene isolated from
Tripterygium wilfordii
.
Cancer Lett, 112(1):113-117.
[33] Sun, Y., Li, S., Song, H., 2006. Extraction of ferulic acid from
Angelica sinensis with supercritical CO
2
.
Nat Prod Res, 20(9):835-841.
[34] van der Vliet, A., O'Neill, C.A., Cross, C.E., 1999. Determination of low-molecular-mass antioxidant concentrations in human respiratory tract lining fluids.
Am J Physiol, 276(2 Pt 1):289-296.
[35] Wang, G., Liu, J.S., Lin, B.B., 2009. Two new furanoid norditerpenes from
Dioscorea bulbifera
.
Chem Pharm Bull, 57(6):625-627.
[36] Wang, J., Ji, L., Liu, H., 2010. Study of the hepatotoxicity induced by
Dioscorea bulbifera L. rhizome in mice.
Biosci Trends, 4(2):79-85.
[37] Wang, J., Liang, Q., Ji, L., 2011. Gender-related difference in liver injury induced by
Dioscorea bulbifera L. rhizome in mice.
Hum Exp Toxicol, 30(9):1333-1341.
[38] Wang, J., Ji, L., Branford-White, C.J., 2012. Antitumor activity of
Dioscorea bulbifera L. rhizome
in vivo
.
Fitoterapia, 83(2):388-394.
[39] Yilmaz, H.R., Turkoz, Y., Yuksel, E., 2006. An investigation of antioxidant enzymes activities in liver of
Cyprinus carpio taken from different stations in the Karakaya Dam Lake.
Int J Food Sci Tech, 1(1):1-6.
[40] You, Y., Park, J., Yoon, H.G., 2009. Stimulatory effects of ferulic acid on endurance exercise capacity in mice.
Biosci Biotechnol Biochem, 73(6):1392-1397.
[41] Zelko, I.N., Mariani, T.J., Folz, R.J., 2002. Superoxide dismutase multigene family: a comparison of the CuZn-SOD (SOD1), Mn-SOD (SOD2), and EC-SOD (SOD3) gene structures, evolution, and expression.
Free Radical Biol Med, 33(3):337-349.




Open peer comments: Debate/Discuss/Question/Opinion
<1>