1. Introduction
Soil and water contaminated with heavy metals have serious consequences for the environment and human health (Tanhan et al.,
2007). Copper (Cu) and zinc (Zn) are important environmental pollutants owing to their elevated concentrations and bioavailability in natural ecosystems (Kopponen et al.,
2001). Cu and Zn contamination originates from mining, smelting, textile, microelectronic and hydrometallurgical industries, sewage sludge, fossil-fuel combustion, and agricultural sources (including fertilizers, manures, pesticides, and fungicides containing Cu or Zn) (Kopponen et al.,
2001; Paschke et al.,
2006). Cu and Zn are also essential elements in plant nutrition and development. However, the presence of Cu and Zn at high concentrations induces severe phytotoxicity, resulting in the reduction of plant growth, mineral nutrient uptake, and photosynthetic activity (Salemaa and Monni,
2003; Yan et al.,
2006; Borghi et al.,
2007; Disante et al.,
2010).
Phytoremediation is the process of using trees, shrubs, and grasses to de-contaminate soil and water, and is considered to be a promising, cost-effective solution to heavy-metal pollution (Licht and Isebrands,
2005; Vangronsveld et al.,
2009; Xing et al.,
2012; Lu et al.,
2013). Generally, phytoremediation and especially phytoextraction rely on plants known as hyperaccumulators; however, these are species that often have a low biomass and slow growth rate, leading to a long time periods required for metal removal (Tanhan et al.,
2007). Recently, willows have been shown to be good candidates for remediating metal-contaminated sites because of their rapid growth, large biomass, deep roots, and high accumulation capacity for heavy metals (Kuzovkina and Quigley,
2005; Kuzovkina and Volk,
2009). In the last decade, short-rotation willow coppices have provided an economic and practical solution for remediation as well as for bioenergy production (Rockwood et al.,
2004; Mirck et al.,
2005).
Apart from genetic modifications, increasing the metal bioaccumulation capacity of a plant is primarily accomplished through the artificial selection of
Salix clones; the selection of appropriate species or clones is crucial for achieving successful phytoremediation (Zalesny et al.,
2007; Zalesny and Bauer,
2007a;
2007b). Numerous studies have revealed a remarkable genotypic variability of the genus
Salix in its ability to tolerate/accumulate heavy metals (Wang and Greger,
2004; Dos Santos Utmazian et al.,
2007; Purdy and Smart,
2008; Zhivotovsky et al.,
2010). Most results of willow screening have shown that the tolerance and uptake of heavy metals depend strongly on species and clones (Punshon et al.,
1995; Dos Santos Utmazian et al.,
2007; Zacchini et al.,
2009; Zhivotovsky et al.,
2010).
Salix comprises about 450 species as well as a large number of willow clones produced through breeding projects worldwide, which together provide a large number of plants to be screened for phytoremediation (Kuzovkina et al.,
2004; Kuzovkina and Quigley,
2005). Owing to the high genetic variability in
Salix, the use of clone selection has been proposed to find those best suited for phytoremediation purposes (Zalesny et al.,
2007; Zalesny and Bauer,
2007b).
Hydroponic screening is the primary method for evaluating the tolerance and accumulation potential of heavy metals in
Salix (Watson et al.,
1999;
2003). Many studies have investigated the metal accumulation potential of
Salix to identify prospective phytoremediation species/clones through carefully controlled hydroponic screenings (Punshon et al.,
1995; Watson et al.,
2003; Kuzovkina et al.,
2004; Wang and Greger,
2004; Dos Santos Utmazian et al.,
2007; Purdy and Smart,
2008; Mleczek et al.,
2010; Zhivotovsky et al.,
2010). As a rapid screening technology, hydroponic screening reduces the time and space required in comparison with other methods, and is especially suitable for high-throughput screening to evaluate large amounts of plant materials for phytoremediation (Dos Santos Utmazian et al.,
2007). However, short-term exposure to metals in a hydroponic system results in large quantities of metal accumulating in the roots, especially metals with low mobility in plants, as this leads to low translocation of metals from the roots to the shoots (Cosio et al.,
2006; Dos Santos Utmazian et al.,
2007; Zhivotovsky et al.,
2010). Consequently, metal accumulation potentials are easily underestimated in willows. Zhivotovsky et al. (
2011) concluded that willows may need more time to adapt to a specific environmental contaminant in order to improve its transport to above-ground plant tissues.
To the best of our knowledge, most recently published research has focused on the toxicities of Cu and Zn in Salicaceae rather than their accumulation capacity, especially for Cu (Punshon et al.,
1995; Borghi et al.,
2007;
2008; Baud and Pezeshki,
2011), and therefore, the Cu accumulation capacity of willows is still poorly understood. In fact, the metal tolerance and accumulation capacity of a plant depend on several factors, including plant species, genotype, growth stage, treatment concentration, and exposure time (Monni et al.,
2000; Weng et al.,
2005; Maxted et al.,
2007; Dauthieu et al.,
2009; Colzi et al.,
2011).
In previous studies, the willows investigated were limited to European and North American species.
Salix has a high genetic diversity worldwide, and China has a rich willow resource with 275 native species that have not yet been explored or utilized (Karp et al.,
2011). In this study, the willow clones tested belong to Asian species, and they are commonly planted in China for biomass production on account of high biomass productivity and fast growth rates. In order to overcome the drawbacks of hydroponic screening, we selected relatively large seedlings and used prolonged exposure time to Cu and Zn to evaluate their uptake and accumulation potential in 12 willow clones.
2. Materials and methods
2.1. Plant materials and growth
The 12 willow clones (Table
1) used in this study were obtained from the Jiangsu Academy of Forestry, Nanjing (33°31′ N, 118°47′ E), China. They were maintained in a nursery in the Experimental Station of Agriculture of Zhejiang University, Hangzhou (30°16′ N, 120°12′ E), China. One-year-old hardwood stems from each clone were collected from the nursery and prepared in uniformly sized 10-cm long cuttings, which were mounted with styrofoam in plastic buckets containing a nutrient solution. Plants were pre-grown in the nutrient solution for 42 d. The composition of the nutrient solution was as follows: 1 mmol/L Ca(NO
3)
2, 1.25 mmol/L KNO
3, 0.5 mmol/L MgSO
4, 0.5 mmol/L NH
4H
2PO
4, and the micronutrients including 25 μmol/L FeEDTA, 23.1 μmol/L H
3BO
3, 0.4 μmol/L ZnCl
2, 0.18 μmol/L CuCl
2, 4.57 μmol/L MnCl
2, and 0.06 μmol/L Na
2MoO
4, as described by Watson et al. (
2003). When the cuttings were well developed, they were used to evaluate metal tolerance and accumulation potential. In this study, according to our preliminary investigation on metal exposure concentrations, treatments containing 50 μmol/L Cu or 50 μmol/L Zn (added as CuSO
4 or ZnSO
4) were performed. The controls were maintained in the above nutrient solution.
Table 1
Salix clones tested
| Entry No. |
Clone |
Species/hybrid |
| 1 |
SB7 |
Salix babylolica
|
| 2 |
SM16 |
S. matsudana
|
| 3 |
SM30 |
S. matsudana
|
| 4 |
SI63 |
S. integra
|
| 5 |
J9-6 |
S. integra×S. suchowensis
|
| 6 |
SI646 |
S. integra
|
| 7 |
J795 |
S. matsudana×S. alba
|
| 8 |
J844 |
S. baylonica×S. alba
|
| 9 |
J1011 |
S. baylonica×S. alba
|
| 10 |
J1052 |
S. suchowensis×S. leucopithecia
|
| 11 |
SI102-1 |
S. integra
|
| 12 |
SI102-2 |
S. integra
|
All treatments were performed in triplicate, and each treatment had three seedlings. The pH of the nutrient solution was maintained at 5.5–6.0 using H
2SO
4/NaOH. Solutions were aerated using a pump throughout the experiment and replaced weekly. The experiment was a randomized complete block design, and all plastic buckets were randomly arranged inside a greenhouse. The greenhouse conditions were 30/18 °C day/night temperature and a relative humidity of 70%–80%.
2.2. Plant samples and analysis
Cu and Zn exposure lasted for 50 d. After this time, all plants were harvested and separated into leaves, stems, and roots. The roots were rinsed carefully in 10 mmol/L ethylenediaminetetraacetic acid (EDTA)-Na
2 for 15 min to remove the adsorbed metals from root surfaces, and then washed in double-deionized water. Plant materials were dried at 70 °C for 48 h in a forced-air oven, and the dry weights (DWs) were recorded. The dried material was ground to a fine powder in a mill (Retsch 2M 200, Germany), and approximately 0.2 g of the plant sample was digested using a HNO
3:HClO
4 solution (4:1, v/v) in a Hot Block (ED36, LabTech, Germany). Cu and Zn concentrations in the extracts were measured using atomic absorption spectrometry (Analyst 800, PerkinElmer Inc., USA). Chemical analyses were validated by blanks and reference materials.
2.3. Calculation and statistical analysis
Following Deng et al. (
2007), tolerance indexes (TIs) were calculated based on the dry shoot biomass (including leaf and stem) or root biomass treated relative to the controls, i.e., TI=DW
treated plant/DW
control. The metal concentration in the shoots (combined leaves and stems) was calculated according to the following equation: metal concentration in shoots=(stem biomass×stem metal concentration+leaf biomass×leaf metal concentration)/(stem biomass+leaf biomass). Following Deng et al. (
2007), the translocation factors (TFs) of metals from roots to leaves or shoots were calculated as follow: TF=leaf or shoot metal concentration/root metal concentration. According to Dos Santos Utmazian et al. (
2007), the total metal content (μg/plant) in the shoots was calculated by multiplying the shoot biomass (DW) by the metal concentration in the shoots.
The main purpose of the experiments was to examine clonal differences under Cu or Zn exposure. To analyze the differences in the treatments and clones simultaneously, treatment and clone effects were determined by two-way analyses of variance (ANOVA) followed by Tukey-HSD test (
P<0.05). All statistics were performed using SPSS software (Version 16.0, SPSS Inc., Chicago, USA).
3. Results
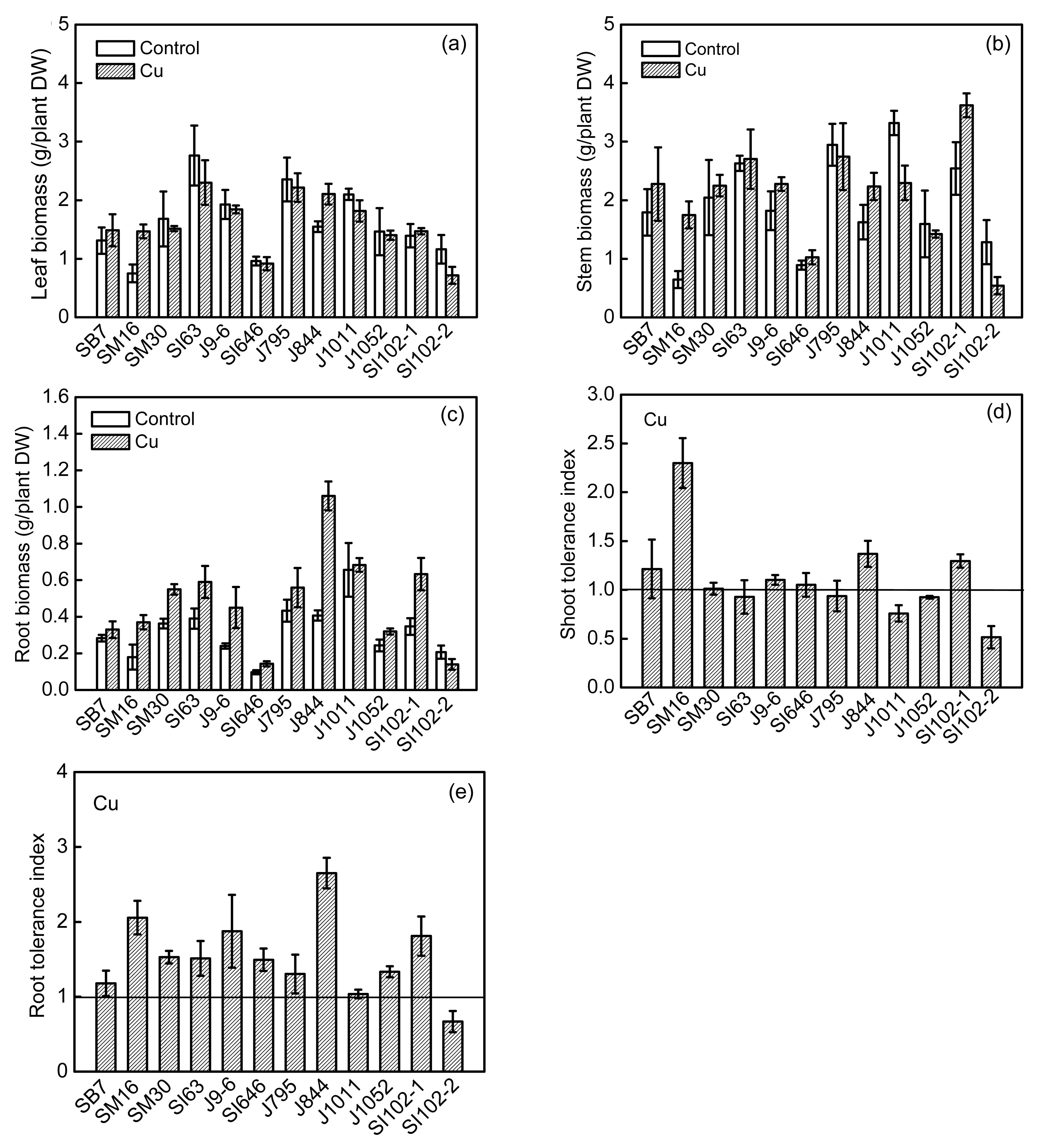
3.1. Biomass production and tolerance
Biomass accumulation and the relative growth of shoots and roots are important parameters for characterizing metal tolerance.
The variation of the DW of tissue biomass for clones exposed to Cu treatments (Fig.
1) shows that Cu depressed leaf biomass production in five clones (clones SM30, SI63, J1011, SI102-2, and J795). In contrast, Cu treatment had little effect on the leaf biomasses of clones J9-6, SI646, J1052, and SI102-1, and it enhanced the leaf biomasses of clones SB7, SM16, and J844 by 13%, 94%, and 35%, respectively, compared with the control (Fig.
1a). Fig.
1b shows the enhanced stem biomasses of eight of the treated clones compared with the control. Cu excess, however, reduced the stem biomasses of clones J795, J1011, J1052, and SI102-2. Other than for clone SI102-2, Cu stimulated root biomass growths of all clones compared with the controls (Fig.
1c). Figs.
1a and 1b also show that the leaves and stems were more sensitive to Cu than the roots.
Fig.1
Dry biomasses of leaves (a), stems (b) and roots (c), and tolerance indexes of shoots (d) and roots (e) of 12 willow clones with 50 μmol/L Cu treatment in hydroponic culture for 50 d
Tolerance indexes were estimated as metal-treated shoot or root dry weight over untreated one. The values represented mean±standard error (SE), with n=3
TIs were expressed as the relative weights of shoots and roots. The Cu TIs based on shoot biomass ranged from 0.51 to 2.29, and seven out of twelve clones showed high tolerance to Cu (TI>1), while clones J1011 and SI102 were more sensitive to Cu (Fig.
1d). The Cu TIs based on root biomass ranged from 0.66 to 2.05 among clones, and root TIs of eleven clones were >1 (Fig.
1e).
The growth response of the clones to excess Zn is also presented in Fig.
2. The foliar biomass accumulation of most clones decreased in response to excess Zn. In contrast, Zn treatment had little inhibitory effect on the leaf biomasses of clones SM16 and J844. However, Zn treatment increased the leaf biomasses of clones SB7 and SI63 compared with the control (Fig.
2a). Exposure to excess Zn reduced the stem biomasses of six clones, and clone SI102-1 showed the largest reduction (56%) compared with the control. In contrast, the stem biomass accumulation of clone SI63, J844, or J1052 was not affected by Zn excess, and Zn exposure significantly increased the stem biomasses of clones SB7 and SM30 compared with the control (Fig.
2b). Excess Zn strongly inhibited the root biomass productions of seven clones; clones SM16, SI646, J844, and J1052 were not affected. However, clone SB7 had a larger root biomass than the control plants (Fig.
2c).
Fig.2
Dry biomasses of leaves (a), stems (b) and roots (c), and tolerance indexes of shoots (d) and roots (e) of 12 willow clones with 50 μmol/L Zn treatment in hydroponic culture for 50 d
Tolerance indexes were estimated as metal-treated shoot or root dry weight over untreated one. The values represented mean±standard error (SE), with n=3
The TIs to Zn treatment were calculated as for Cu. The Zn TIs based on shoot biomass ranged from 0.57 to 1.67 (Fig.
2d), and the root-based TIs ranged from 0.43 to 1.32 among clones (Fig.
2e). Considering both shoot- and root-based TIs (Figs.
2d and 2e), clones SB7 and J844 showed high tolerances to Zn (TIs>1), and seven clones showed medium tolerances (0.7
3.2. Cu and Zn accumulation
The accumulation and translocation of Cu in willow clones are shown in Fig. 3. The Cu concentration in the roots always exceeded that in the stems or leaves. The Cu concentration in the leaves ranged from 391.1 (J844) to 660.83 μg/g DW (SB7) under Cu exposure, varying by a factor of 1.6 among the clones (Fig. 3a). The Cu concentration in the stems ranged from 417.5 (SM30) to 786.2 μg/g DW (SI102-2) under conditions of excess Cu, varying by a factor of 1.8 among clones (Fig. 3b). The Cu concentration in the roots ranged from 2065.3 μg/g DW for SI102-1 to 2876.2 μg/g DW for SB7 under Cu treatment, varying by a factor of 1.4 (Fig. 3c). The TFs for Cu in the leaves ranged from 0.17 to 0.27 under Cu exposure (Fig. 3d), and similarly, the TFs for Cu in the shoots (leaves and stems) ranged from 0.21 to 0.27 (Fig. 3e). The TFs for Zn from the roots to the stems and leaves were much greater than those for Cu.
Fig.3
Variations of Cu concentrations in leaves (a), stems (b) and roots (c), and translocation factors in leaves (d) and shoots (e) of 12 willow clones with 50 μmol/L Cu treatment in hydroponic culture for 50 d
Translocation factors were shown as leaf or shoot metal concentration divided by root metal concentration. The values represented mean±standard error (SE), with n=3
The accumulation and translocation of Zn in willow clones are shown in Fig. 4. The Zn concentration was higher in the roots than in the leaves and stems for eight clones; however, four clones showed higher Zn concentrations in the leaves than in the stems and roots, i.e., clones J795, J844, J1011, and J1052 under the Zn treatment. The Zn concentration in the leaves ranged from 607.1 (SI646) to 1218.8 μg/g DW (clone J1011) under exposure to Zn, varying by a factor of 2 among clones (Fig. 4a). The Zn concentration in the stems ranged from 258.8 μg/g DW for J844 to 1455.5 μg/g DW for SM30, varying by a factor of 5.6 (Fig. 4b). The Zn concentration in the roots ranged from 575.8 (J844) to 2052.8 μg/g DW (SM30), varying by a factor of 3.6 (Fig. 4c).
Fig.4
Variation of Zn concentrations in leaves (a), stems (b) and roots (c), and translocation factors in leaves (d) and shoots (e) of 12 willow clones with 50 μmol/L Zn treatment in hydroponic culture for 50 d
Translocation factors were shown as leaf or shoot metal concentration divided by root metal concentration. The values represented mean±standard error (SE), with n=3
The leaf TFs for Zn ranged from 0.4 to 1.81, and leaf TFs for six clones were close to or >1 under Zn exposure (Fig. 4d). The shoot TFs for Zn ranged from 0.39 to 1.12, and there were four clones with TFs near to or larger than 1 (Fig. 4e). Combining leaf TFs with shoot TFs, clones J795, J844, J1011, and J1052 showed higher Zn transfer capacities than the other clones.
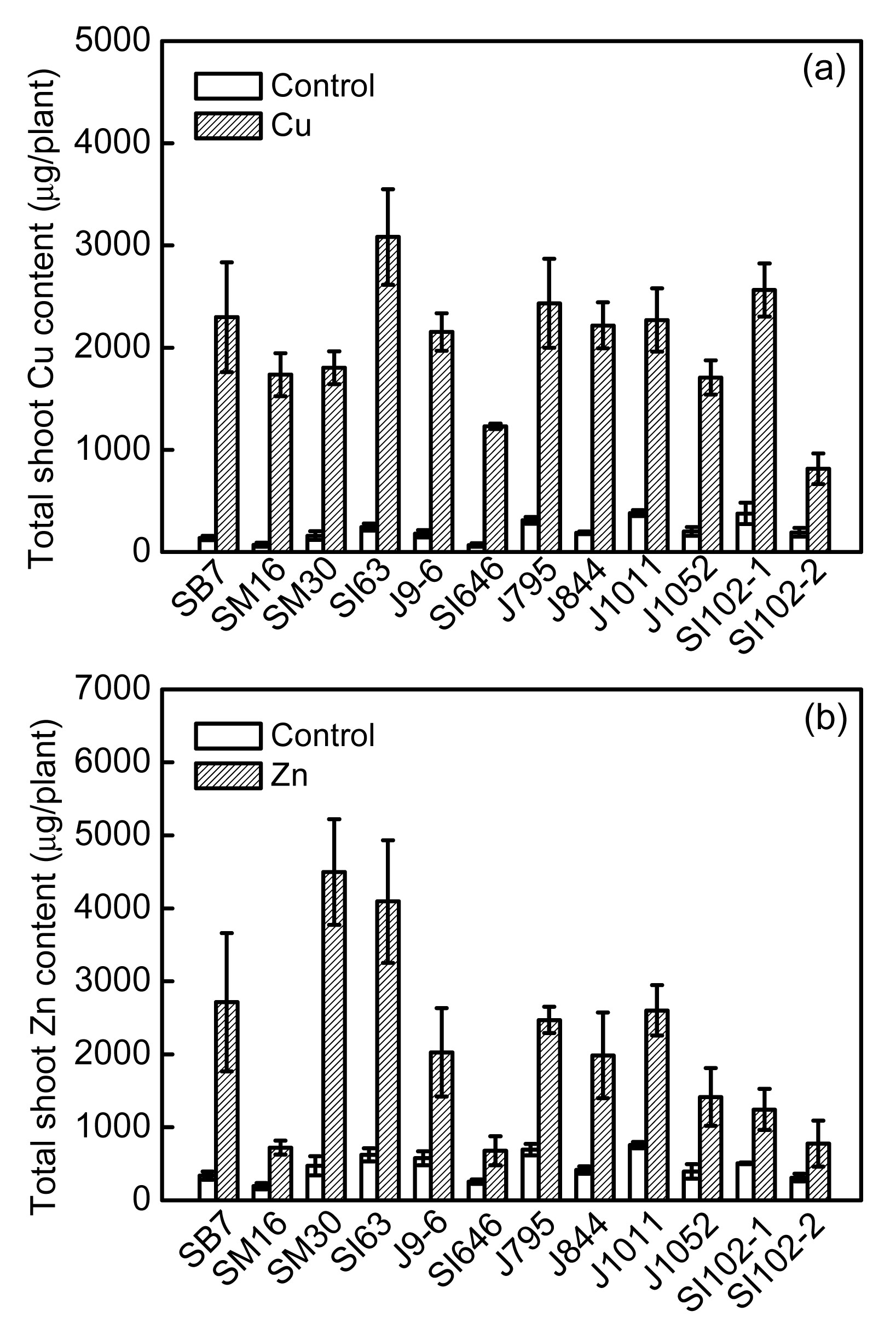
3.3. Cu and Zn phytoextraction potentials
The metal contents in shoot tissues (combined stems and leaves) were calculated by multiplying the DWs of plant parts by the metal concentrations in the corresponding parts. The Cu and Zn contents of the shoots varied significantly among clones. Fig. 5a shows Cu contents in the shoots, ranging from 814.5 (SI102-2) to 3083.0 μg/plant (SI63) under Cu treatment, thus displaying a 3.8-fold variation among all clones. The shoot Zn contents varied between 677.6 (SI646) and 4497.7 μg/plant (SM30) under Zn excess, showing a 6.6-fold variation (Fig. 5b).
Fig.5
Total phytoextraction Cu (a) or Zn (b) in the shoots of 12 willow clones with 50 μmol/L Cu or Zn treatments in hydroponic culture for 50 d
The values represented mean±standard error (SE), with n=3
4. Discussion
4.1. Variation in plant growth and tolerance
No symptoms of necrosis or chlorosis were found in the leaves, suggesting that the levels of Cu and Zn in this study were not severely phytotoxic to the willow clones. The results showed wide variations in biomass accumulation in response to Cu and Zn, ranging from causing an increase, decrease, to having no significant effect. These results are consistent with previous findings, which illustrated high genetic variability in the tolerance of the Salicaceae family toward heavy metals (Punshon et al., 1995; Dos Santos Utmazian et al., 2007; Zacchini et al., 2009; Zhivotovsky et al., 2010).
Our data showed that the response to Cu exposure depended on the plant tissues. Cu treatment inhibited leaf biomass production in most clones, whereas it stimulated stem biomass production. Other than for SI102-2, Cu was shown to stimulate root growth in the treated willow clones compared with controls. These results are different from those reported by Punshon et al. (1995). We calculated the TI based on the relative growths of the shoots and roots, and the results suggest that the clones tested have a relatively high tolerance to Cu, except for clones J1011 and SI102-2, and both the shoot and root TIs of seven clones were found to be >1. Indeed, as an essential microelement, the present study showed that Cu enhanced plant growth under the current Cu treatment concentration. Therefore, under the current experimental conditions, willows can be described as Cu-tolerant plants, similar to the results reported by Borghi et al. (2007) on poplars.
The effect on growth in response to Zn was also highly dependent on the species and clones. Excess Zn resulted in a decrease in the biomass of most clones. Unlike Cu, the roots were more sensitive to toxic Zn concentrations than the shoots. Clone SB7 had the highest tolerance to Zn, and in fact, it strongly stimulated the growth of this clone, indicating a hypertolerance to the 50 μmol/L Zn treatment.
Our results also indicated that Zn toxicity was higher compared with Cu toxicity; similar results have also been reported for some Mediterranean crops by López-Gonzálvez et al. (1999). Clones SB7, SM16, and J844 showed a significantly higher tolerance to both Cu and Zn than the other clones, while clones J1011 and SI102-2 were found to be more sensitive to both Cu and Zn than other clones.
4.2. Variation in Cu and Zn accumulation
The present data suggest that there are small variations in Cu accumulation among clones, and in contrast, a comparatively wide variation in Zn concentration. These results are in agreement with previous observations, confirming the genetic variation of metal accumulation in Salix (Punshon et al., 1995; Dos Santos Utmazian et al., 2007; Purdy and Smart, 2008; Zacchini et al., 2009; Zhivotovsky et al., 2010).
The values presented in Fig. 3 indicate that clones accumulated higher percentages of Cu in the roots than in aerial parts, which is in agreement with previous reports (Ali et al., 2003; Kuzovkina et al., 2004; Borghi et al., 2007; 2008; Castiglione et al., 2009). The distribution pattern of Cu in plant parts was ranked in the order of roots>stems>leaves. Our results showed relatively high levels of Cu in the above-ground parts, especially in the aerial stems; similar results were reported by Punshon and Dickinson (1997). To the best of our knowledge, however, our results indicate that the Cu accumulation ability of the clones investigated here is far higher than those reported in other studies of the Salicaceae family. Comparing this with previous results, Punshon et al. (1995) found that the Cu concentration for willows was in the range of 9.5–63.0 μg/g DW in the leaves and 10–47 μg/g DW in the stems; according to Ali et al. (2003), the Cu concentration of Salix acmophylla was 10.8–126.3 μg/g DW in leaves, 25.3–203.7 μg/g DW in stems, and 24.8–624.4 μg/g DW in roots. Punshon and Dickinson (1997) reported that Salix fragilis accumulated Cu >300 μg/g DW in leaves, 25–150 μg/g DW in stems, and 300–1200 μg/g DW in roots. However, under some experimental conditions, relatively high Cu concentrations in aerial parts of the Salicaceae family were also recorded. For example, Turchi et al. (2012) reported that Cu accumulation in Populus alba ranged from 128.87 to 312.92 μg/g DW in the leaves, and from 1401.66 to 1598.92 μg/g DW in the roots; Komárek et al. (2010) found high Cu concentrations (377 μg/g DW) in the leaves of poplars grown in soil; Castiglione et al. (2009) also showed that P. alba accumulated Cu >400 μg/g DW in leaves, >100 μg/g DW in stems, and >500 μg/g DW in roots. Therefore, owing to the different experimental conditions, these results are difficult to compare.
There seems to be considerable variation in reports on the uptake and accumulation of Cu. For some plants, although Cu is a micronutrient, many studies do not show high translocation from the roots to above-ground organs. For the Cu concentration in the roots, our results were similar to other reports on poplar, but in the shoots, our results were far higher than those in the poplars reported by Borghi et al. (2007; 2008). Under hydroponic conditions, Cu noticeably accumulated in the roots and there was almost no transport of Cu from the roots to the shoots (Borghi et al., 2007; 2008). Borghi et al. (2007) found that the Cu concentration in the leaves and stems was only about 10 μg/g DW, while the root Cu concentration reached 2000–4000 μg/g DW; similarly, Borghi et al. (2008) noted that the average Cu concentration was 5.9–8.8 μg/g DW in the leaves and stems, whereas the root Cu concentration reached 6×105–1.4×106 μg/g DW. Guerra et al. (2009) found the Cu concentration in the leaves of Populus deltoides to be close to 50 μg/g DW, while the Cu concentration in the roots ranged from 5000 to 6000 μg/g DW. Indeed, these studies on poplar using the hydroponic system evaluated Cu toxicity tolerance rather than Cu accumulation potential. These findings suggest that almost no Cu accumulation occurred in the shoot tissues compared with the roots, i.e., that Cu absorption was largely confined to the roots, and therefore, the findings may lead to an underestimate of the Cu accumulation capacity in aerial parts compared with experiments using soils (Castiglione et al., 2009; Komárek et al., 2010; Turchi et al., 2012).
However, other studies have indicated considerable Cu accumulation and translocation to above-ground organs of woody plants, showing higher accumulation of Cu in aerial parts. For example, Monni et al. (2000) reported that the shrub, Empetrum nigrum, accumulated 690 μg/g Cu in leaves and 3038 μg/g in stems; Sahi et al. (2007) found that the shrub, Sesbania drummondi, accumulated 1282 μg Cu/g DW in shoots, and 27 440 μg/g DW in roots. These different results may be due to the different species/genotypes, treatment concentrations, culture conditions, and exposure time.
In fact, Cu accumulation ability depends on many factors. For example, Cu accumulation increased with seedling age (Monni et al., 2000); the optimum growth stage of the plants enhanced the maximum Cu uptake on account of a much higher transpiration rate, which enhances metal uptake (Arduini et al., 2006; Mleczek et al., 2010; Weng et al., 2005). Colzi et al. (2012) reported that Cu accumulation in the apoplast and symplast of roots was higher in one-month-old plants than in two-month-old plants. Culture conditions and exposure time can also affect metal uptake in plants. In particular, short-term exposure in hydroponic systems may not be sufficient for metal accumulation to occur in aerial parts of willows, leading to higher metal concentrations in the roots (Cosio et al., 2006; Dos Santos Utmazian et al., 2007).
Consequently, under the experimental conditions in this study, genotype, seedling size, growth stage, treatment concentration, and exposure time might enhance Cu accumulation in the aerial parts of the willow clones. Relatively large seedlings and a good growth stage increased metabolic activity, leading to absorption and transport of larger amounts of Cu (Weng et al., 2005; Arduini et al., 2006; Dauthieu et al., 2009).
The TF of plants is an important parameter for evaluating the potential for phytoremediation by harvesting above-ground parts of the plants (Deng et al., 2007). The TF of Cu was lower than that of Zn, ranging from 0.17 to 0.27 in leaves and from 0.14 to 0.47 in shoots. Low values of leaf and shoot TFs in the Cu treatments suggest that all clones had a poorer ability to translocate Cu from the roots to the leaves/shoots (including stems). This low mobility of Cu in the Salicaceae family has also been confirmed in previous studies (Punshon et al., 1995; Kuzovkina et al., 2004; Castiglione et al., 2009). The low translocation of Cu to the shoots is probably due to the roots having a strong affinity for Cu (Sahi et al., 2007), as Cu is confined to the root surface or is adsorbed by the apoplast (Kuzovkina et al., 2004); further, Cu binds to the carboxylic groups of pectins and polygalacturonic acids in the roots (Borghi et al., 2007).
For most willow clones, the largest Zn accumulation was found in the roots, followed by the leaves and stems, but in some clones, higher percentages of Zn were found in the leaves rather than in the roots. Under the experimental conditions of this study, Zn concentrations in the leaves were close to or larger than 1000 μg/g DW; clone SM30 accumulated the highest concentration of Zn in the stems, reaching 1455 μg/g DW. The findings demonstrated that all of the clones had a high phytoextraction potential for Zn. The Zn accumulation capacity in this study is similar to those in other reports. For example, the largest Zn concentration (3180 μg/g DW) in leaves was detected in a Salix smithiana clone grown in soil containing 955 mg/kg Zn (Dos Santos Utmazian et al., 2007). Wieshammer et al. (2007) reported large Zn concentrations (3300 μg/g DW) in the leaves of Salix× smithiana. Castiglione et al. (2009) found that the Zn concentrations of P. alba were >2500 μg/g DW in leaves, >700 μg/g DW in stems, and close to 1200 μg/g DW in roots.
High values of leaf and shoot TFs for the Zn treatments suggest that most willow clones possessed a good ability to translocate Zn from their roots to their stems/leaves. Clones J795, J844, J1011, and J1052 had a greater tendency for Zn translocation from their roots to their shoots than other clones, demonstrated by their shoot TFs being >1. Although clone SM30 accumulated 1455 μg Zn/g DW in above-ground parts, which exceeded the Zn hyperaccumulator threshold, the shoot TFs did not exceed 1.0; hence, clone SM30 did not behave as a Zn hyperaccumulator. Previous studies have shown that Zn is easily translocated to the leaves of willow (Dos Santos Utmazian et al., 2007).
In this study, nearly all willow clones exhibited the largest amount of Cu accumulation in their roots. In contrast, the concentration of Zn in aerial parts was higher than that of Cu. Cu was poorly translocated to the aerial parts of the willow clones, and thus, Zn is more mobile than Cu within a plant, this result being similar to the finding in Chromolaena odorata (Tanhan et al., 2007). As shown in Figs. 3 and 4, most clones tested did not show a common uptake pattern for the two metals; these results indicated absorptions of Cu and Zn by willows via independent uptake mechanisms, as has been demonstrated for other plants (Ait Ali et al., 2004).
4.3. Variation in phytoextraction potential
Tissue concentrations should not be used to evaluate the phytoremediation ability of species, because they do not take into consideration the whole plant biomass and metal concentration in the growth media (Ait Ali et al., 2002). The phytoextraction efficiency of plants depends on the amount of heavy metals accumulated in the above-ground plant parts and the plant yields (Deng et al., 2007). The total uptake of metals by the plants was calculated by multiplying the metal concentrations in each plant organ by the corresponding dry mass (Dos Santos Utmazian et al., 2007).
Despite low TFs of Cu in willow clones, the present results show that willows have relatively high phytoextraction potential for Cu owing to its high biomass production. Clone SI63 showed the highest phytoextraction potential for Cu (3083.00 μg/plant DW in shoots), while clone SI102-2 showed the lowest shoot Cu content (814.59 μg/plant DW); the shoot Cu content in SI63 was about 4-fold higher than that in SI102-2 under the same Cu treatment. This was because clone SI63 had a larger biomass than other clones and a relatively high Cu accumulation capacity in its aerial parts. In contrast, despite the high concentration in its shoots, SI102-2 showed a poor phytoextraction capacity for Cu owing to its low biomass. Most clones produced more root biomass and a large proportion of Cu retention in roots exposed to excess Cu; hence, these results indicate that willows could be a good candidate for the phytostabilization of Cu.
The highest Zn content (4497.7 μg/plant DW) in the shoots was recorded in clone SM30 exposed to excess Zn, while the lowest Zn content (677.6 μg/plant DW) was found in clone SI102-2; the variation between clones SM30 and SI646 was 6.6-fold. Based on the total Zn content in its shoots, clone SM30 had the highest Zn phytoextraction potential, while SI646 had low phytoextraction potential, owing to the poor growth performance of SI646 under the Zn treatment. Hence, these results also confirm that the selection and improvement of willow clones for phytoextraction should focus on biomass production.
5. Conclusions
The results of this study demonstrated the variation among willow clones with respect to Cu/Zn tolerance and accumulation capacity. The willow clones investigated were less sensitive to Cu than to Zn, showing high resistance to 50 μmol/L of Cu. Although Cu translocation to above-ground plant parts was low, willows showed good Cu phytoextraction potential owing to the large biomass and relatively high Cu concentration in shoots. Zn was more easily translocated to the above-ground tissues and mainly accumulated in the stems, thus showing higher phytoextraction potential than Cu. Therefore, based on relatively high Cu and Zn contents in the above-ground parts, our study showed that most willow clones are good accumulators for Zn and Cu. This study also highlights that the genetic variations in Salix can be exploited to improve their phytoremediation efficiency by selection of the most appropriate species/clones.
Acknowledgements
We thank Ms. Hua-xian ZHANG, from the Analysis Center of Agrobiology and Environmental Science of Zhejiang University, China, for her excellent technical assistance.
* Project supported by the National Natural Science Foundation of China (No. 31100513), the Key International Cooperative Project of CN-USA from the Ministry of Science and Technology of China (No. 2010DFB 33960), the Key Project of Ningbo Science and Technology Bureau of Zhejiang Province (No. 2012C10003), the Key Project of the Ministry of Science and Technology of China (No. 2012BAC17B02), and the Fundamental Research Funds for the Central Universities, ChinaCompliance with ethics guidelines Wei-dong YANG, Yu-yan WANG, Feng-liang ZHAO, Zhe-li DING, Xin-cheng ZHANG, Zhi-qiang ZHU, and Xiao-e YANG declare that they have no conflict of interest.
References
[1] Ait Ali, N., Bernal, M.P., Ater, M., 2002. Tolerance and bioaccumulation of copper in
Phragmites australis and
Zea mays
.
Plant Soil, 239(1):103-111.


[2] Ait Ali, N., Bernal, M.P., Ater, M., 2004. Tolerance and bioaccumulation of cadmium by
Phragmites australis grown in the presence of elevated concentrations of cadmium, copper, and zinc.
Aquat Bot, 80(3):163-176.
[3] Ali, M.B., Vajpayee, P., Tripathi, R.D., 2003. Phytoremediation of lead, nickel, and copper by
Salix acmophylla Boiss.: role of antioxidant enzymes and antioxidant substances.
Bull Environ Contam Toxicol, 70(3):462-469.
[4] Arduini, I., Ercoli, L., Mariotti, M., 2006. Response of miscanthus to toxic cadmium applications during the period of maximum growth.
Environ Exp Bot, 55(1-2):29-40.
[5] Baud, D.R., Pezeshki, S.R., 2011. Root porosity changes in
Salix nigra cuttings in response to copper and ultraviolet-B radiation exposure.
Water Air Soil Pollut, 221(1-4):99-105.
[6] Borghi, M., Tognetti, R., Monteforti, G., 2007. Responses of
Populus×
euramericana (
P. deltoides×
P. nigra) clone
Adda to increasing copper concentrations.
Environ Exp Bot, 61(1):66-73.
[7] Borghi, M., Tognetti, R., Monteforti, G., 2008. Responses of two poplar species (
Populus alba and
Populus×canadensis) to high copper concentrations.
Environ Exp Bot, 62(3):290-299.
[8] Castiglione, S., Todeschini, V., Franchin, C., 2009. Clonal differences in survival capacity, copper and zinc accumulation, and correlation with leaf polyamine levels in poplar: a large-scale field trial on heavily polluted soil.
Environ Pollut, 157(7):2108-2117.
[9] Colzi, I., Doumett, S., Del Bubba, M., 2011. On the role of the cell wall in the phenomenon of copper tolerance in
Silene paradoxa L.
Environ Exp Bot, 72(1):77-83.
[10] Cosio, C., Vollenweider, P., Keller, C., 2006. Localization and effects of cadmium in leaves of a cadmium-tolerant willow (
Salix viminalis L.): I. Macrolocalization and phytotoxic effects of cadmium.
Environ Exp Bot, 58(1-3):64-74.
[11] Dauthieu, M., Denaix, L., Nguyen, C., 2009. Cadmium uptake and distribution in
Arabidopsis thaliana exposed to low chronic concentrations depends on plant growth.
Plant Soil, 322(1-2):239-249.
[12] Deng, D.M., Shu, W.S., Zhang, J., 2007. Zinc and cadmium accumulation and tolerance in populations of
Sedum alfredii
.
Environ Pollut, 147(2):381-386.
[13] Disante, K.B., Fuentes, D., Cortina, J., 2010. Sensitivity to zinc of Mediterranean woody species important for restoration.
Sci Total Environ, 408(10):2216-2225.
[14] Dos Santos Utmazian, M.N., Wieshammer, G., Vega, R., 2007. Hydroponic screening for metal resistance and accumulation of cadmium and zinc in twenty clones of willows and poplars.
Environ Pollut, 148(1):155-165.
[15] Guerra, F., Duplessis, S., Kohler, A., 2009. Gene expression analysis of
Populus deltoides roots subjected to copper stress.
Environ Exp Bot, 67(2):335-344.
[16] Karp, A., Hanley, S.J., Trybush, S.O., 2011. Genetic improvement of willow for bioenergy and biofuelsfree access.
J Integr Plant Biol, 53(2):151-165.
[17] Komrek, M., Vaněk, A., Mrnka, L., 2010. Potential and drawbacks of EDDS-enhanced phytoextraction of copper from contaminated soils.
Environ Pollut, 158(7):2428-2438.
[18] Kopponen, P., Utriainen, M., Lukkari, K., 2001. Clonal differences in copper and zinc tolerance of birch in metal-supplemented soils.
Environ Pollut, 112(1):89-97.
[19] Kuzovkina, Y.A., Quigley, M.F., 2005. Willows beyond wetlands: uses of
Salix L. species for environmental projects.
Water Air Soil Pollut, 162(1-4):183-204.
[20] Kuzovkina, Y.A., Volk, T.A., 2009. The characterization of willow (
Salix L.) varieties for use in ecological engineering applications: co-ordination of structure, function and autecology.
Ecol Eng, 35(8):1178-1189.
[21] Kuzovkina, Y.A., Knee, M., Quigley, M.F., 2004. Cadmium and copper uptake and translocation in five willow (
Salix L.) species.
Int J Phytoremediat, 6(3):269-287.
[22] Licht, L.A., Isebrands, J.G., 2005. Linking phytoremediated pollutant removal to biomass economic opportunities.
Biomass Bioenerg, 28(2):203-218.
[23] Lpez-Gonzlvez, A., Probanza, A., Galli, V., 1999. Tolerance of some mediterranean crops to copper and zinc: implications in toxic metal clean up.
Chem Ecol, 16(4):297-316.
[24] Lu, L., Tian, S., Yang, X., 2013. Improved cadmium uptake and accumulation in the hyperaccumulator
Sedum alfredii: the impact of citric acid and tartaric acid.
J Zhejiang Univ-Sci B (Biomed & Biotechnol), 14(2):106-114.
[25] Maxted, A.P., Black, C.R., West, H.M., 2007. Phytoextraction of cadmium and zinc by
Salix from soil historically amended with sewage sludge.
Plant Soil, 290(1-2):157-172.
[26] Mirck, J., Isebrands, J.G., Verwijst, T., 2005. Development of short-rotation willow coppice systems for environmental purposes in Sweden.
Biomass Bioenerg, 28(2):219-228.
[27] Mleczek, M., Kaczmarek, Z., Magdziak, Z., 2010. Hydroponic estimation of heavy metal accumulation by different genotypes of
Salix
.
J Environ Sci Health A: Tox Hazard Subst Environ Eng, 45(5):569-578.
[28] Monni, S., Salemaa, M., Millar, N., 2000. The tolerance of
Empetrum nigrum to copper and nickel.
Environ Pollut, 109(2):221-229.
[29] Paschke, M.W., Perry, L.G., Redente, E.F., 2006. Zinc toxicity thresholds for reclamation forb species.
Water Air Soil Pollut, 170(1-4):317-330.
[30] Punshon, T., Dickinson, N.M., 1997. Acclimation of
Salix to metal stress.
New Phytol, 137(2):303-314.
[31] Punshon, T., Lepp, N.W., Dickinson, N.M., 1995. Resistance to copper toxicity in some British willows.
J Geochem Explor, 52(1-2):259-266.
[32] Purdy, J.J., Smart, L.B., 2008. Hydroponic screening of shrub willow (
Salix spp.) for arsenic tolerance and uptake.
Int J Phytoremediat, 10(6):515-528.
[33] Rockwood, D.L., Naidu, C.V., Carter, D.R., 2004. Short-rotation woody crops and phytoremediation: opportunities for agroforestry?.
Agroforest Syst, 61-62(1-3):51-63.
[34] Sahi, S.V., Israr, M., Srivastava, A.K., 2007. Accumulation, speciation and cellular localization of copper in
Sesbania drummondii
.
Chemosphere, 67(11):2257-2266.
[35] Salemaa, M., Monni, S., 2003. Copper resistance of the evergreen dwarf shrub
Arctostaphylos uva-ursi: an experimental exposure.
Environ Pollut, 126(3):435-443.
[36] Tanhan, P., Kruatrachue, M., Pokethitiyook, P., 2007. Uptake and accumulation of cadmium, lead and zinc by Siam weed [
Chromolaena odorata (L.) King & Robinson].
Chemosphere, 68(2):323-329.
[37] Turchi, A., Tamantini, I., Camussi, A.M., 2012. Expression of a metallothionein A1 gene of
Pisum sativum in white poplar enhances tolerance and accumulation of zinc and copper.
Plant Sci, 183:50-56.
[38] Vangronsveld, J., Herzig, R., Weyens, N., 2009. Phytoremediation of contaminated soils and groundwater: lessons from the field.
Environ Sci Pollut Res, 16(7):765-794.
[39] Wang, Y., Greger, M., 2004. Clonal differences in mercury tolerance, accumulation, and distribution in willow.
J Environ Qual, 33(5):1779-1785.
[40] Watson, C., Pulford, I.D., Riddell-Black, D., 1999. Heavy metal toxicity responses of two willow (
Salix) varieties grown hydroponically: development of a tolerance screening test.
Environ Geochem Health, 21(4):359-364.
[41] Watson, C., Pulford, I.D., Riddell-Black, D., 2003. Development of a hydroponic screening technique to assess heavy metal resistance in willow (
Salix).
Int J Phytoremediat, 5(4):333-349.
[42] Weng, G., Wu, L., Wang, Z., 2005. Copper uptake by four
Elsholtzia ecotypes supplied with varying levels of copper in solution culture.
Environ Int, 31(6):880-884.
[43] Wieshammer, G., Unterbrunner, R., Garca, T.B., 2007. Phytoextraction of Cd and Zn from agricultural soils by
Salix ssp. and intercropping of
Salix caprea and
Arabidopsis halleri
.
Plant Soil, 298(1-2):255-264.
[44] Xing, Y., Peng, H., Li, X., 2012. Extraction and isolation of the salidroside-type metabolite from zinc (Zn) and cadmium (Cd) hyperaccumulator
Sedum alfredii Hance.
J Zhejiang Univ-Sci B (Biomed & Biotechnol), 13(10):839-845.
[45] Yan, Y.P., He, J.Y., Zhu, C., 2006. Accumulation of copper in brown rice and effect of copper on rice growth and grain yield in different rice cultivars.
Chemosphere, 65(10):1690-1696.
[46] Zacchini, M., Pietrini, F., Mugnozza, G.S., 2009. Metal tolerance, accumulation and translocation in poplar and willow clones treated with cadmium in hydroponics.
Water Air Soil Pollut, 197(1-4):23-34.
[47] Zalesny, J.A., Zalesny, R.S., Wiese, A.H., 2007. Choosing tree genotypes for phytoremediation of landfill leachate using phyto-recurrent selection.
Int J Phytoremediat, 9(6):513-530.
[48] Zalesny, R.S., Bauer, E.O., 2007. Evaluation of
Populus and
Salix continuously irrigated with landfill leachate I. genotype-specific elemental phytoremediation.
Int J Phytoremediat, 9(4):281-306.
[49] Zalesny, R.S., Bauer, E.O., 2007. Selecting and utilizing
Populus and
Salix for landfill covers: implications for leachate irrigation.
Int J Phytoremediat, 9(6):497-511.
[50] Zhivotovsky, O.P., Kuzovkina, J.A., Schulthess, C.P., 2010. Hydroponic screening of willows (
Salix L.) for lead tolerance and accumulation.
Int J Phytoremediat, 13(1):75-94.




Open peer comments: Debate/Discuss/Question/Opinion
<1>