1. Introduction
Among the polysaccharides produced by bacteria, β-glucans consisting of a backbone of β-1,3-D-glucose units with various lengths of β-1,6 side chains have the notable physiological effect of immunological activation (Miura et al.,
1996). Immunologists have found the β-glucan receptors, dectin-1 and complement receptor 3, on the surface of innate immune cells (Brown and Gordon,
2001; Vetvicka et al.,
2007). There are many application studies associated with β-glucan immune activation, e.g., tumoricidal activity, prevention of infection, and damage reduction of radiation exposure (Thompson et al.,
2010). β-Glucans are also found to decrease the levels of serum cholesterol and liver low-density lipoproteins, leading to lowering of atherosclerosis and cardiovascular disease hazards (Barsanti et al.,
2011). Salecan, produced by the strain
Agrobacterium sp. ZX09, is a novel water-soluble β-glucan-like polysaccharide. It consists of the following repeating units: →3)-β-
D-Glcp-(1→3)-[β-
D-Glcp-(1→3)-β-
D-Glcp-(1→3)]
3-α-
D-Glcp-(1→3)-α-
D-Glcp-(1→ (Xiu et al.,
2010). This special structure of Salecan is water-soluble and has potential biological activities owing to its β-1,3-glucosides. It reduces the acute hepatic injury in mice induced by alcohol (Chen et al.,
2011) and carbon tetrachloride (Chen et al.,
2012). It has a potential to become a hydrophilic laxative to cure constipation (Zhou et al.,
2013). It can also possibly be used in the food industry as a thickening agent due to its excellent rheological properties (Xiu et al.,
2011). Therefore, increasing the yield of Salecan has a vital significance for its industrial application.
It is well known in fermentation by aerobic organisms that the supply of O
2 usually becomes a serious limitation because of its low solubility, especially during high-cell-density fermentation or when fermentation media are highly viscous.
Vitreoscilla, an obligate aerobic bacterium (Pringsheim,
1951), survives and grows in an O
2-poor environment owing to its synthesis of hemoglobin (VHb). VHb binds O
2 at low extracellular concentrations and delivers it to the terminal respiratory oxidase, thus enhancing respiration (Park et al.,
2002). The
vgb gene from
Vitreoscilla has been successfully engineered into various organisms to improve cell density, protein synthesis, and metabolic productivity (Khosla and Bailey,
1988; Frey and Kallio,
2006; Su et al.,
2010; Frey et al.,
2011). The
vgb gene is generally integrated into the genome of the host organisms to form stable inheritance, but this technique is usually limited to the genome-sequenced organisms.
In this study, a broad-host-range vector pCM158 (Marx and Lidstrom,
2002), hosting
vgb gene, was demonstrated to persist in Salecan-producing
Agrobacterium sp. ZX09 without antibiotic selection pressure and to improve the respiration rate, invertase activity, and Salecan production.
2. Materials and methods
2.1. Strains, plasmids, media, and culture conditions
Escherichia coli strain DH5α was used as the host for bacterial transformation and plasmid construction. The plasmid pCM158 was used as
vgb expression vector, and the
vgb gene was derived from pUC19-vgb. Salecan-producing bacterium,
Agrobacterium sp. ZX09, was isolated from a soil sample from the ocean coast of Shandong, China (Xiu et al.,
2010). The culture medium for screening transformation and producing Salecan was Htm composed of NaH
2PO
4 1 g, KNO
3 3 g, CaCl
2 0.07 g, MgCl
2 0.2 g, FeSO
4·7H
2O 0.0125 g, MnSO
4 0.003 g, ZnCl
2 0.0075 g, sucrose 30 g, and H
2O 1000 ml, pH 7.2 (Xiu et al.,
2010). The strain hosting pCM158-vgb plasmid was maintained on Htm agar medium by adding 50 μg/ml kanamycin. A single colony on an agar plate was first inoculated in seed medium (Htm) and aerobically incubated at 30 °C for 24 h with shaking at 250 r/min. A 1% inoculum was added to a 250-ml flask containing 50 ml of medium. The flask culture was incubated at 30 °C for 72 h at 250 r/min and the yield of Salecan was measured after cultivation for 12, 24, 36, 48, and 72 h.
2.2. Construction of pCM158-vgb plasmid and transformation
The VHb coding gene
vgb was first cloned from the plasmid pUC19-vgb (Khosla and Bailey,
1988) by polymerase chain reaction (PCR) with the two primers of vgb-F (AAGCTTACAGGACGCTGGGGT) and vgb-R (CAATATTTGTCCCAAGTTT). The PCR products were then cloned into pMD18-T vector to form pMD18-T-vgb, and sequenced to be correct without mutations. Last, the
vgb gene fragment was cut down from the pMD18-T-vgb by restriction endonucleases
SphI and
EcoRI, and cloned into the corresponding sites of pCM158 plasmid to form pCM158-vgb. Electrocompetent cells were prepared in super optimal broth (SOB) medium. Once the optical density at 600 nm (OD
600) of the culture reached about 0.5 to 0.6, the cells were placed on ice and washed three times with ice-cold 10% glycerol. About 100 ng of pCM158-vgb and pCM158 DNA were added to 100 μl of the competent cells and the resulting mixtures were electroporated in 0.1-cm-gap cuvettes at 2.0 kV, 25 μF, 200 Ω in a Gene Pulser electroporation apparatus (BioRad). Cells were resuspended in 1 ml SOB medium. The cells were allowed to recover at 30 °C for 2 h, and then plated on Htm agar plates with 50 μg/ml kanamycin. Strains of
Agrobacterium sp. ZX09 hosting the plasmids of pCM158-vgb and pCM158 were respectively designated VHb
+ and VHb
−. Plasmid stability was studied by inoculation of the strain into Htm medium without antibiotics and incubating at 28 °C with shaking (Guo et al.,
2010). After 48 h of cultivation, the same amounts of cells were sprayed onto six solid medium plates: half of the plates lacked antibiotics and half contained antibiotics. Plasmid stability was indicated by the ratio of the colony-forming units (CFUs) formed on the plates with antibiotics to the ones formed on the plates without antibiotics.
2.3. CO-difference spectral analysis of VHb
The
vgb gene expression was determined by carbon monoxide (CO) difference spectra of VHb (Liu and Webster,
1974). Cells of VHb
+ and VHb
− were harvested from 100 ml Htm medium after cultivation, then washed with potassium phosphate buffer (KH
2PO
4 5.44 g/L, Na
2HPO
4·12H
2O 43 g/L; pH 7.2), disrupted by grinding in liquid nitrogen, and resuspended in 5 ml of potassium phosphate buffer. The cell debris was separated by centrifugation at 15 000×
g for 5 min. The liquid samples were exposed to CO for 10 min and then used for CO-difference spectral analysis by PowerWave HT Microplate Spectrophotometer (BioTek Instruments, Inc.).
2.4. Analytical methods
The viscosity of Salecan was determined by a NDJ-1 rotational viscometer (Shanghai Changji Instruments Co., Ltd., China) to indicate the relative yield of the sugar. The productivity of exopolysaccharide was expressed in terms of the weight after ethanol precipitation, collected by centrifugation, and dried under reduced pressure (Xiu et al.,
2010).
The respiratory rate was determined by inoculating 1 ml of the seed medium into a 1-L flask bottle containing 250 ml Htm medium. The bottle was sealed by a cap, and incubated at 30 °C on a rotary shaker at 150 r/min. The decrease in dissolved O
2 was recorded online by an optical O
2 sensor VisiFerm DO Arc 120 (Hamilton Company) inserting through the cap.
Invertase activities were measured by DNS colorimetry (Miller,
1959). Cells of VHb
+ and VHb
− were separated from 100 ml culture broth by centrifugation, grinded in liquid nitrogen, and resuspended with 2 ml sodium phosphate buffer (0.2 mol/L NaH
2PO
4 93.5 ml, 0.2 mol/L Na
2HPO
4 6.5 ml; pH 5.7). A 500-μl sample of the cell suspension was mixed with 500 μl of 0.1 g/ml sucrose solution. After 10 min, the hydrolysis was stopped by immersion of the reaction mixture in a boiling water bath for 5 min. Glucose generated from sucrose hydrolysis by invertase was measured by DNS colorimetry. Invertase activities were expressed as milligrams of glucose per minute per milligram of protein (mg/(min∙mg)).
3. Results
3.1. Maintenance of pCM158-vgb and its expression in Agrobacterium sp. ZX09 without antibiotic selection
The
vgb gene was cloned into the broad-host-range plasmid pCM158 to form pCM158-vgb (Fig.
1), and then pCM158-vgb was successfully introduced into
Agrobacterium sp. ZX09 by electroporation. The plasmid persisted well in
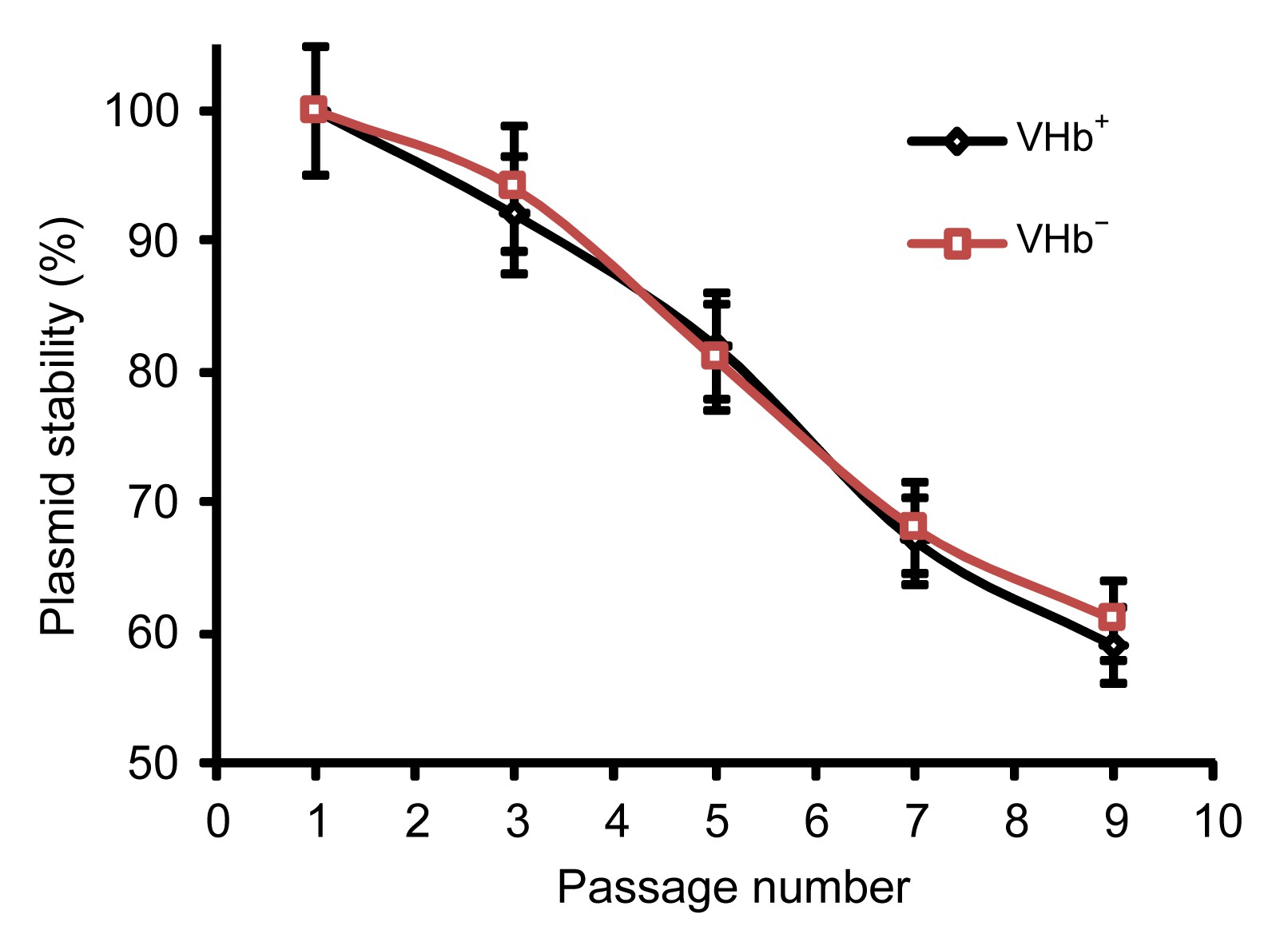
Agrobacterium sp. ZX09. Under no antibiotic selection, there was a negligible loss of the plasmid for one passage of two days cultivation (Fig.
2), but after five passages of cultivation the loss of the plasmid was apparent. Plasmid loss is not easy as commonly thought. Velappan et al. (
2007) have shown that even some incompatible plasmids could persist together in host for several passages without antibiotic selection. Persistence of plasmids in host without antibiotic selection was crucial for their practical usage in fermentation; the use of antibiotics would dramatically increase the cost of fermentation, and the waste containing the antibiotics discharged into the environment would pose serious health problems. Foreign genes were usually integrated into the genome of the host for stable propagation. However, it needs double crossover and clear genetic background of the host. The process is time-consuming, especially for non-model organisms. Another shortcoming is that the genomic-integrated foreign gene is single-copied, thus limiting the gene product. On the contrary, plasmids can overcome these shortcomings. They are multi-copied and easy to handle without the genetic background of the host. In this experiment, we showed that the broad-host-range plasmid pCM158 can persist well in
Agrobacterium sp. ZX09 without antibiotic selection at the end of cultivation and should have practical usage for the increase of product yield.
Fig.1
Schematic map of plasmid pCM158-vgb
Fig.2
pCM158 and pCM158-vgb maintenance in Agrobacterium sp. ZX09 without antibiotic selection
One passage was defined as 48-h cultivation in Htm medium without antibiotics. Plasmid stability was indicated by the ratio of the CFUs formed on the plates with antibiotics to those formed on the plates without antibiotics. Results were averages (±standard deviation) of triplicate experiments
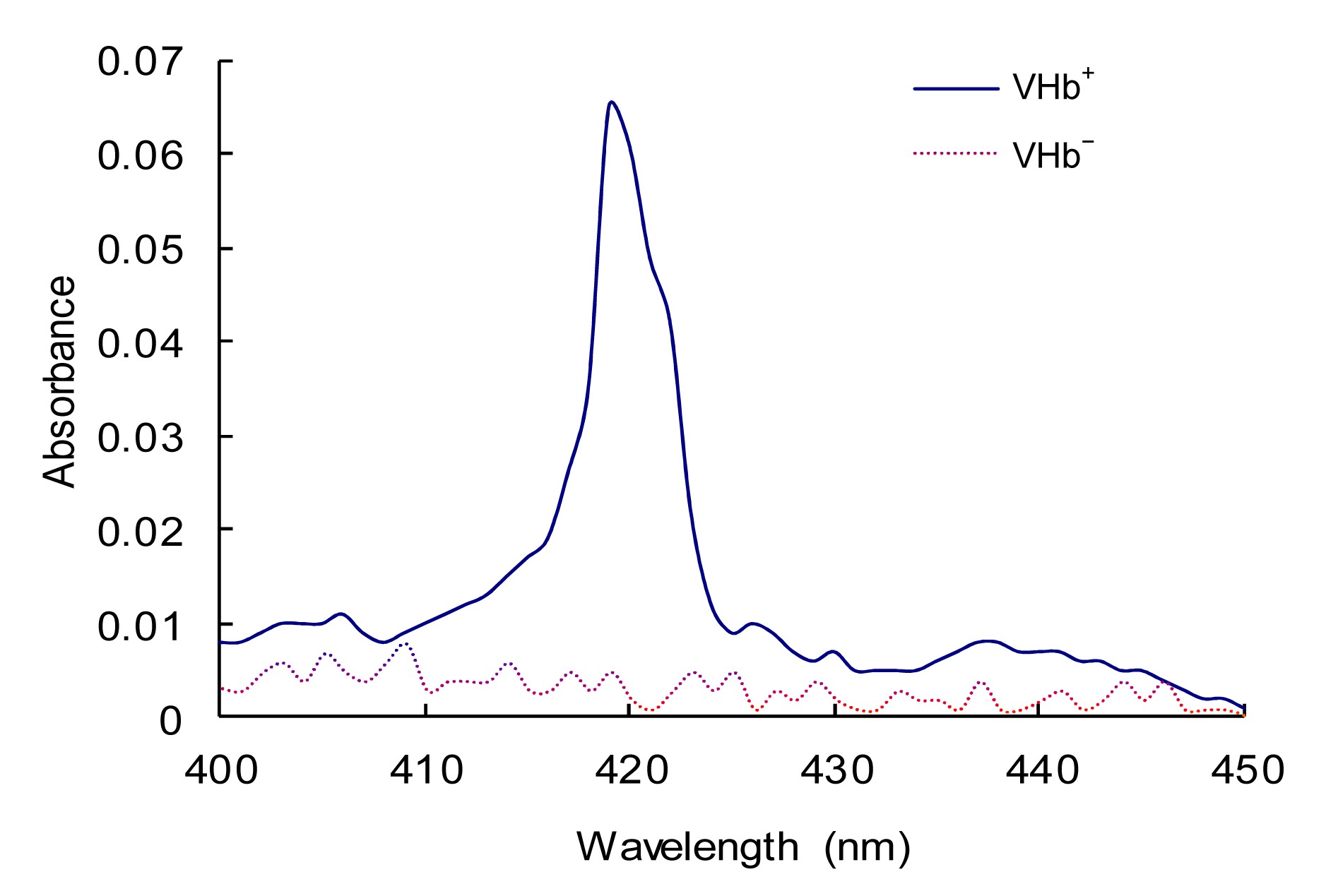
The expression of VHb was demonstrated by CO-difference spectrum owing to its binding to CO. The absorbance spectrum of the solution is subtracted from the spectrum after saturation with CO. Bubbling CO shifted the absorption maxima to 419 nm, which are characteristic of the VHb-CO complex. There was an absorption peak at 419 nm in
Agrobacterium sp. ZX09 hosting pCM158-vgb (VHb
+), but it was absent in the strain hosting the empty vector pCM158 (VHb
−) after bubbling CO (Fig.
3). Cells of VHb
+ and VHb
− were collected for analysis after 2-d culturing in Htm medium without adding antibiotics. These results showed that VHb was stably expressed and further indicated that the plasmid could persist in
Agrobacterium sp. ZX09 without antibiotic selection.
Fig.3
CO-difference spectral analysis of VHb+ and VHb−
Cells of VHb+ and VHb− were collected for analysis after 2-d culturing in Htm medium without adding antibiotics
3.2. VHb increased Salecan production
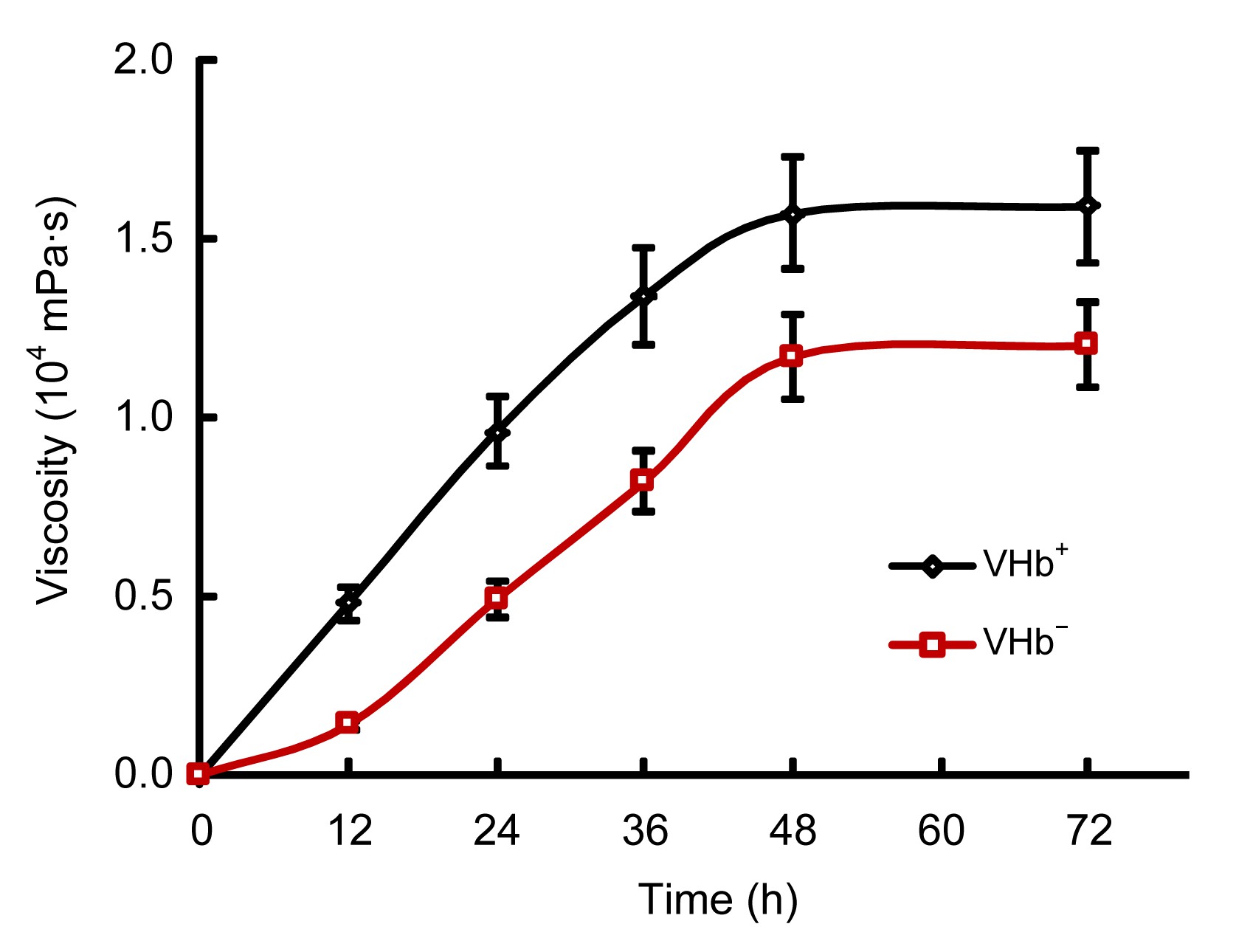
The viscosity yields of the two strains were monitored at 12 h intervals for 72 h (Fig.
4). The viscosity of VHb
+ was apparently higher than that of VHb
−, indicating the higher Salecan production by VHb
+. At the end of fermentation, viscosity yield of VHb
+ was about 33% higher than that of VHb
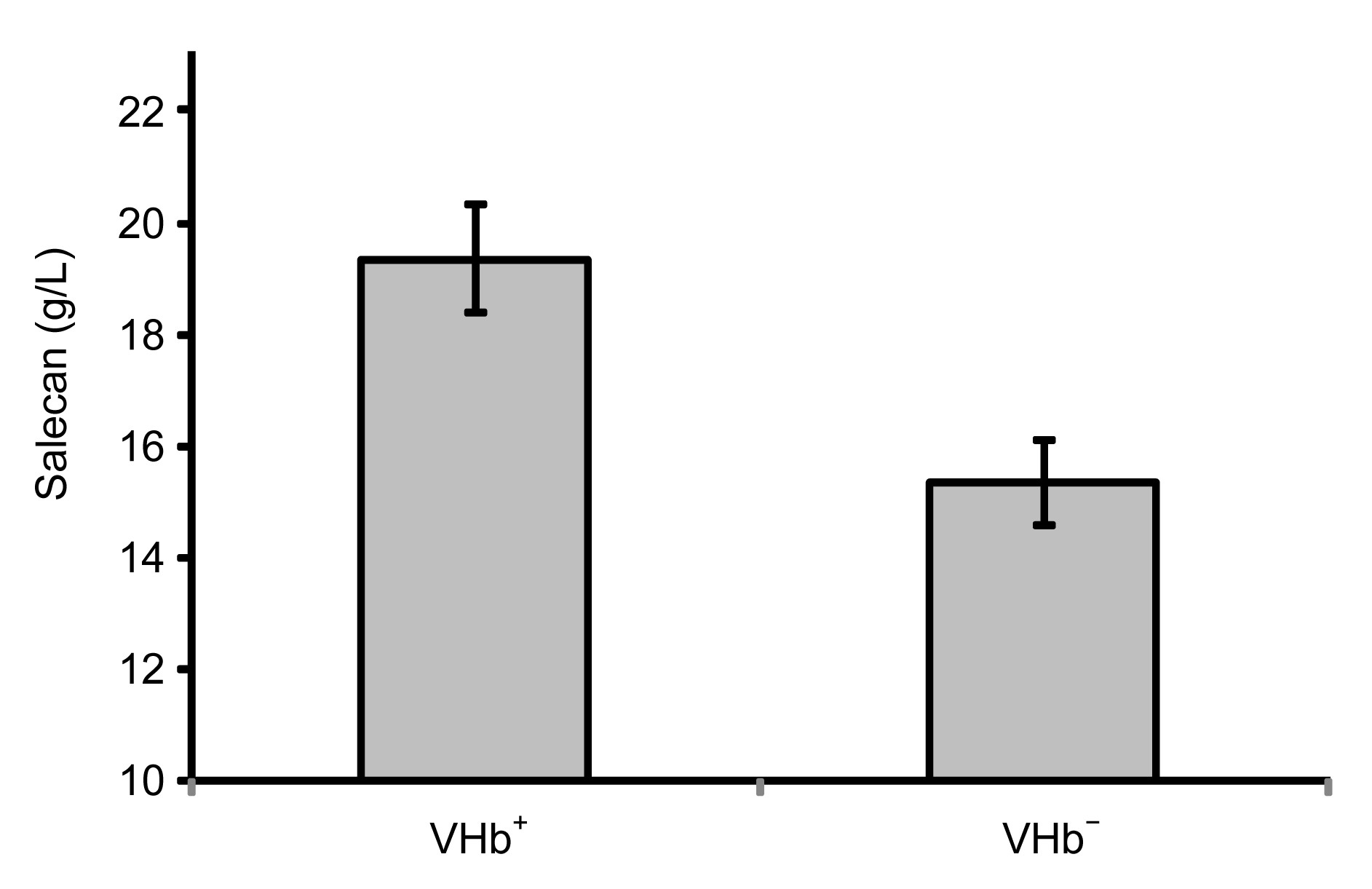
−. The obvious viscosity difference was manifested even at the first 12 h of the fermentation process. The Salecan production of VHb
+ was about 30% higher than that of VHb
− (Fig.
5). Polymer synthesis is a high-energy and O
2 demanding process, whereas polymer in fermentation liquid inhibits O
2 transfer on account of its high viscosity. Therefore, polymer fermentation should be very sensitive to O
2 supply.
Fig.4
Viscosity yields of VHb+ and VHb−
The two strains were monitored at 12 h intervals for 72 h on a rotary shaker at 250 r/min. Results were averages (±standard deviation) of triplicate experiments
Fig.5
Exopolysaccharide productivities of VHb+ and VHb−
The fermentation was performed on a rotary shaker at 250 r/min for 48 h. Results are averages (±standard deviation) of triplicate experiments
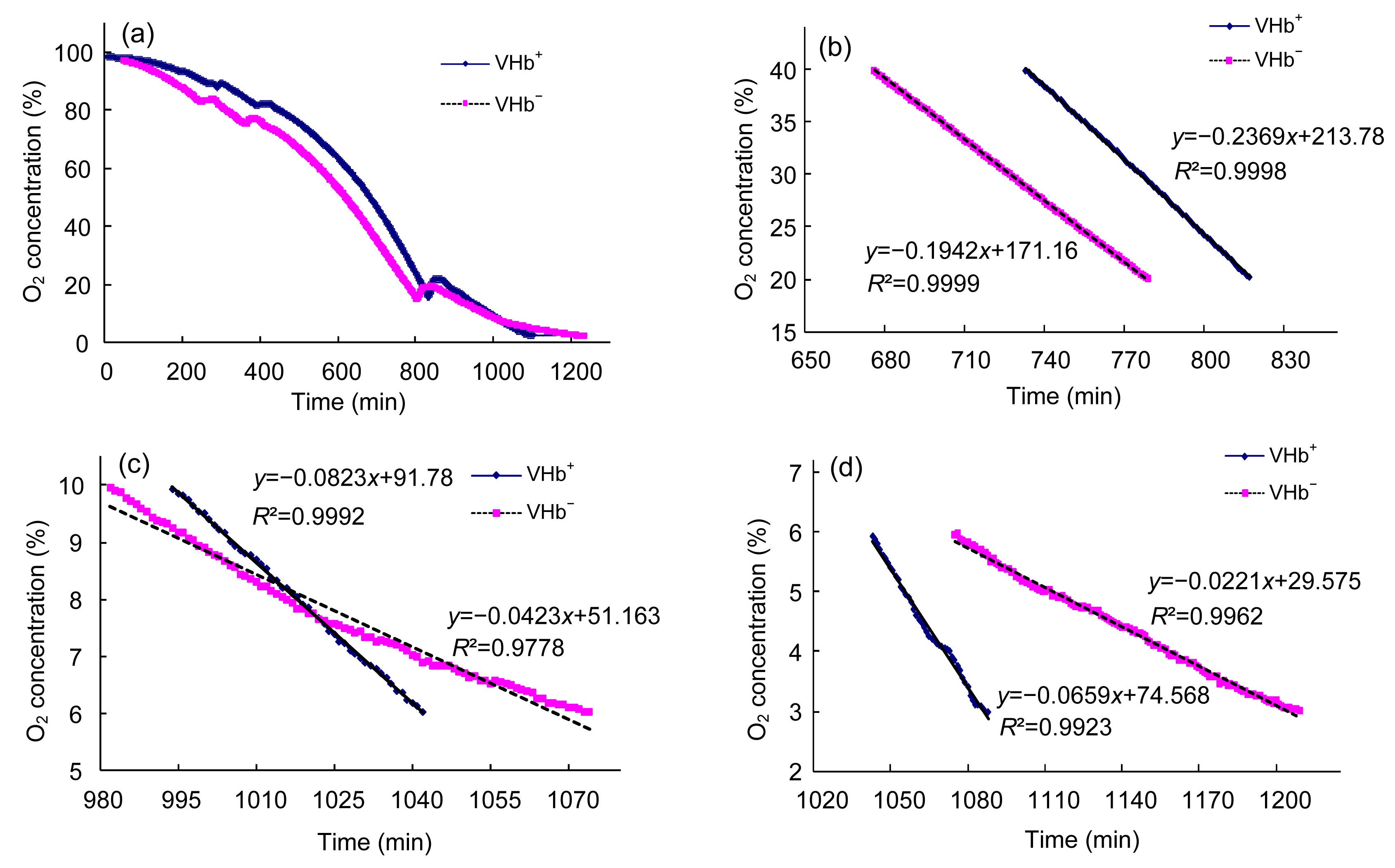
3.3. VHb increased respiration rate
The O
2 concentration in the liquid was monitored online in a sealed bottle. In order to set off the effect of cell concentration, the same amounts of VHb
+ and VHb
− were inoculated, and at the end of the experiment, the OD
600 values of the cells were measured to be 0.048 and 0.046, respectively, for VHb
+ and VHb
−, a difference of less than 5%. In general, the two curves had a similar pattern of O
2 decreasing rate (Fig.
6a), but at different O
2 concentrations the consumption rates varied. The lower the O
2 concentration was, the VHb
+ consumed O
2 at a higher rate. At the interval of 20%–40%, 6%–10%, and 3%–6% air saturated O
2 concentration, the relative respiration rates of VHb
+ were 0.236, 0.082, and 0.065, respectively, while the values for VHb
− were 0.194, 0.042, and 0.022 (Figs.
6b–6d). Both VHb
+ and VHb
− dropped their respiration rates with the decrease of O
2 concentration, but with different dropping rates. VHb
+ consumed O
2 faster than VHb
−. At the three O
2 concentration intervals, the respiration rates of VHb
+ were 1.2, 1.9, and 3.1-fold that of VHb
−, respectively. While at the interval of 40%–100% air saturated O
2 concentration, the difference of respiration rate between VHb
+ and VHb
− was not obvious (data not shown). The growth limiting O
2 concentrations for bacteria are usually far below air saturated O
2 concentration. Above the growth limiting O
2 concentrations, the terminal oxidase is saturated by O
2 and the reaction will not increase with the rise of O
2 concentrations. This explains why at high O
2 concentration of 20%–40% the respiration rates of VHb
+ and VHb
− were similar; at very low O
2 concentration, VHb played a significant role in facilitating its host respiration. The localization of VHb was determined to be concentrated near the cell membrane (Ramandeep et al.,
2001), and further study showed that VHb could bind the subunit I of cytochrome bo and provide O
2 directly to the terminal oxidase (Park et al.,
2002), thus providing O
2 directly to the terminal oxidase.
Fig.6
Respiration rates of VHb+ and VHb−
(a) In general, VHb+ and VHb− had a similar pattern of O2 decreasing rate. The O2 concentration in the liquid was monitored online in a sealed bottle. In order to set off the effect of cell concentration, the same amounts of VHb+ and VHb− were inoculated. (b–d) Respiration rates at 20%–40% (b), 6%–10% (c), and 3%–6% (d) air saturated O2 concentration interval. These formulae indicate the linear regression of the data. The coefficients indicate the change rate of O2 concentration (y) with respect to time (x). R
2, the statistic parameter, measures the goodness of model fitting, with the value of 1 as perfect fitting
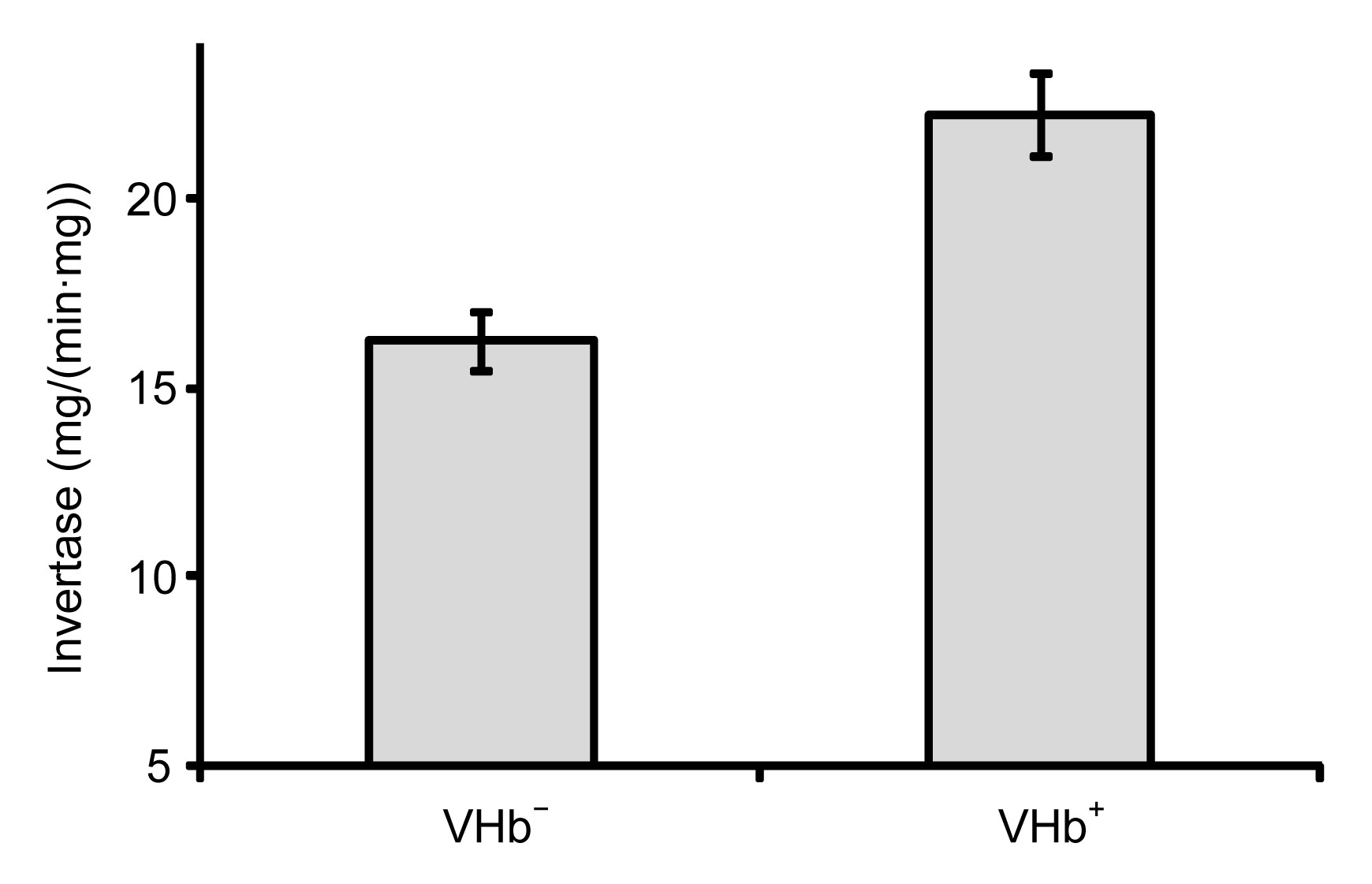
3.4. VHb increased cellular invertase activity
The increase in respiration rate and product yield means more rapid carbon drainage and conversion. Since sucrose was the sole carbon source for
Agrobacterium sp. ZX09, we measured the activity of the sucrose hydrolase enzyme invertase. The cellular invertase activity in VHb
+ increased 37% compared with VHb
− strain (Fig.
7). It was reported that over-expression of invertase in plants increased oxygen consumption (Bologa et al.,
2003). Therefore, the increase in respiration of VHb
+ was probably caused in part by the increase in invertase activity. The mechanism for the increase in invertase activity was unclear, though there were similar reports that the expression of invertase was suppressed under low O
2 concentration (Zeng et al.,
1999; Bologa et al.,
2003).
Fig.7
Invertase activities of VHb+ and VHb−
Invertase activity was expressed as milligram of glucose per minute per milligram of protein (mg/(min∙mg)). Results are averages (±standard deviation) of triplicate experiments
4. Conclusions
In order to overcome the O
2 limiting during Salecan fermentation, we cloned the
Vitreoscilla hemoglobin gene,
vgb, into a broad-host-range plasmid pCM158 to form pCM158-vgb. The plasmid could persist in the host
Agrobacterium sp. ZX09 for at least one passage without antibiotic selection. The engineered strain
Agrobacterium sp. ZX09 increased Salecan yield by 30% without antibiotic selection pressure. The benefit for the approach is its ease of handling and usefulness for field fermentation.
Acknowledgements
We would like to thank Mary E. LIDSTROM from the University of Washington, USA for providing the plasmid pCM158.
* Project supported by the Fundamental Research Funds for the Central Universities of China (No. 30920130121013) and the National Natural Science Foundation of China (No. 31300111)Compliance with ethics guidelines Yun-mei CHEN, Hai-yang XU, Yang WANG, Jian-fa ZHANG, and Shi-ming WANG declare that they have no conflict of interest.References
[1] Barsanti, L., Passarelli, V., Evangelista, V., 2011. Chemistry, physico-chemistry and applications linked to biological activities of β-glucans.
Nat Prod Rep, 28(3):457-466.


[2] Bologa, K.L., Fernie, A.R., Leisse, A., 2003. A bypass of sucrose synthase leads to low internal oxygen and impaired metabolic performance in growing potato tubers.
Plant Physiol, 132(4):2058-2072.
[3] Brown, G.D., Gordon, S., 2001. Immune recognition. A new receptor for β-glucans.
Nature, 413(6851):36-37.
[4] Chen, P., Wang, Z., Zeng, L., 2011. A novel soluble β-glucan Salecan protects against acute alcohol-induced hepatotoxicity in mice.
Biosci Biotechnol Biochem, 75(10):1990-1993.
[5] Chen, P., Wang, Z., Zeng, L., 2012. Protective effects of Salecan against carbon tetrachloride-induced acute liver injury in mice.
J Appl Toxicol, 32(10):796-803.
[6] Frey, A.D., Kallio, P.T., 2006. Bacterial hemoglobins and flavohemoglobins: versatile proteins and their impact on microbiology and biotechnology.
FEMS Microbiol Rev, 27(4):525-545.
[7] Frey, A.D., Shepherd, M., Jokipii-Lukkari, S., 2011. The single-domain globin of
Vitreoscilla: augmentation of aerobic metabolism for biotechnological applications.
Advan Microb Physiol, 58:81-139.
[8] Guo, Q., Zhang, W., Ma, L.L., 2010. A food-grade industrial arming yeast expressing β-1,3-1,4-glucanase with enhanced thermal stability.
J Zhejiang Univ-Sci B (Biomed & Biotechnol), 11(1):41-51.
[9] Khosla, C., Bailey, J.E., 1988. The
Vitreoscilla hemoglobin gene: molecular cloning, nucleotide sequence and genetic expression in
Escherichia coli
.
Mol Gen Genet, 214(1):158-161.
[10] Liu, C.Y., Webster, D.A., 1974. Spectral characteristics and interconversions of the reduced oxidized, and oxygenated forms of purified cytochrome o.
J Biol Chem, 249(13):4261-4266.
[11] Marx, C.J., Lidstrom, M.E., 2002. Broad-host-range
cre-lox system for antibiotic marker recycling in Gram-negative bacteria.
Biotechniques, 33(5):1062-1067.
[12] Miller, G.L., 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugar.
Anal Chem, 31(3):426-428.
[13] Miura, N.N., Ohno, N., Aketagawa, J., 1996. Blood clearance of (1→3)-β-D-glucan in MRL
lpr/
lpr mice.
FEMS Immunol Med Microbiol, 13(1):51-57.
[14] Park, K.W., Kim, K.J., Howard, A.J., 2002.
Vitreoscilla hemoglobin binds to subunit I of cytochrome
bo ubiquinol oxidases.
J Biol Chem, 277(36):33334-33337.
[15] Pringsheim, E.G., 1951. The Vitreoscillaceae: a family of colourless, gliding, filamentous organisms.
J Gen Microbiol, 5(1):124-149.
[16] Ramandeep, ., Hwang, K.W., Raje, M., 2001.
Vitreoscilla hemoglobin. Intracellular localization and binding to membranes.
J Biol Chem, 276(27):24781-24789.
[17] Su, Y., Li, X., Liu, Q., 2010. Improved poly-γ-glutamic acid production by chromosomal integration of the
Vitreoscilla hemoglobin gene (
vgb) in
Bacillus subtilis
.
Bioresource Technol, 101(12):4733-4736.
[18] Thompson, I.J., Oyston, P.C., Williamson, D.E., 2010. Potential of the β-glucans to enhance innate resistance to biological agents.
Expert Rev Anti-Infect Ther, 8(3):339-352.
[19] Velappan, N., Sblattero, D., Chasteen, L., 2007. Plasmid incompatibility: more compatible than previously thought?.
Protein Eng Des Sel, 20(7):309-313.
[20] Vetvicka, V., Dvorak, B., Vetvickova, J., 2007. Orally administered marine (1→3)-β-D-glucan Phycarine stimulates both humoral and cellular immunity.
Int J Biol Macromol, 40(4):291-298.
[21] Xiu, A., Kong, Y., Zhou, M., 2010. The chemical and digestive properties of a soluble glucan from
Agrobacterium sp. ZX09.
Carbohyd Polym, 82(3):623-628.
[22] Xiu, A., Zhou, M., Zhu, B., 2011. Rheological properties of Salecan as a new source of thickening agent.
Food Hydrocolloids, 25(7):1719-1725.
[23] Zeng, Y., Wu, Y., Avigne, W.T., 1999. Rapid repression of maize invertases by low oxygen. Invertase/sucrose synthase balance, sugar signaling potential, and seedling survival.
Plant Physiol, 121(2):599-608.
[24] Zhou, M., Jia, P., Chen, J., 2013. Laxative effects of Salecan on normal and two models of experimental constipated mice.
BMC Gastroenterol, 13:52


Open peer comments: Debate/Discuss/Question/Opinion
<1>